オレキシンは1998年に同定された神経ペプチドであり、覚醒の維持や摂食行動の調節など様々な生理的役割を担っている。オレキシンを含有するニューロンは視床下部外側野に局在し、投射先は脳内に広く及んでいる。中でも特に強い投射が、モノアミン及びコリン作動性神経核にみられ、オレキシンはそれらの神経細胞を強く活性化させることが知られている。これらの神経核は、睡眠ー覚醒の制御において重要な役割を果たすとされている。オレキシンは、これらの神経細胞の活性化を介して睡眠ー覚醒の調節を行っていると考えられる。しかし一方、オレキシンニューロン自体の性質に関しては、その同定の困難さから未だに詳細な情報が得られていない。これは、オレキシンニューロンを含む視床下部外側野が多種の神経細胞が混在する領域であり、生組織標本においてオレキシンニューロンの同定が困難なことに起因する。そこで本研究では我々の研究室で作成したオレキシンニューロン特異的にEnhanced Green Fluorescent Protein (EGFP)を発現するOrexin/EGFPトランスジェニックマウスを用いてオレキシンニューロンを同定し、様々な神経伝達物質に対するオレキシンニューロンの応答を明らかにすることを試みた。
材料と方法
雌雄のOrexin/EGFPマウス(2-4週令)を用いて実験を行った。深く麻酔したマウスから脳を素早く取り出し、トリミングした後、ビブラトームを用い、氷冷した塩化コリンベースのHepes-Buffer中で厚さ300 μmのスライス標本を作成した。得られたスライスは塩化ナトリウムベースのHepes-Buffer中に静置し、室温で1時間程度の回復期間をおいた後、実験に用いた。EGFPを発現したオレキシンニューロンは蛍光顕微鏡下で同定し(図1)、

近赤外微分干渉顕微鏡像を視認しながら電気生理学的実験を行った。記録はホールセルパッチクランプ法で行なった。記録電極の抵抗は4-8 MΩ程度のものを用いた。チャンバー内の温度は37℃に設定し、100%酸素でバブリングしたHepes-Bufferを4 ml/minで灌流した。薬液は局所投与法により細胞の近傍100 μmから投与した。
結果と考察
オレキシンニューロンはスライスパッチクランプ条件下で、-60.5±5.9 mVの静止膜電位を持ち、自発発火頻度は5.5±3.9 Hzであった(n=34)。この値は、近傍のニューロンと有意に異なるものではなかった。 次に、オレキシンニューロンに対する入力系を明らかにする目的で、睡眠ー覚醒の制御に重要な役割を果たしていると考えられるモノアミン、コリン系神経伝達物質である、セロトニン(5HT)、ノルエピネフリン(NE)、ヒスタミン(HA)、ドーパミン(DA)、カルバコール(CCh, アセチルコリン受容体アゴニスト)の作用を解析した。その結果、NE、5HTは100%のオレキシンニューロンで過分極応答(5HT n=70,NE n=65)を示した。一方、CChは26.9%のオレキシンニューロンで脱分極、5.9%のオレキシンニューロンで過分極応答を示した(n=152)(図2)。
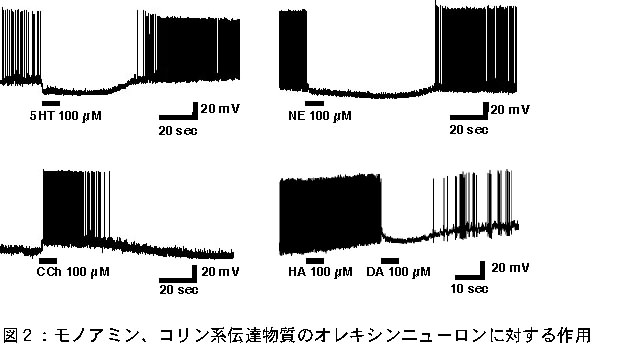
この反応はテトロドトキシン存在下でも確認された。しかし一方、ヒスタミンは全く作用を引き起こさなかった(n=19)。 以上の結果より、オレキシンニューロンはモノアミン、コリン性伝達物質による制御を受けていることが明らかとなった。これらの作用はテトロドトキシンで抑制されなかったことから、オレキシンニューロンにはモノアミン、コリン作動性神経からの直接的なシナプスがあると考えられる。 モノアミン及びコリン系伝達物質を含有するニューロンは、中枢神経系では主に中脳に神経核を作って局在しており覚醒の維持に重要な働きを果たすことが知られている。今回の実験結果は、覚醒を引き起こす神経核から覚醒を引き起こすオレキシンニューロンへの抑制性入力があることを明らかにしている。今後は、この入力系の機能的意義の解明を行う予定である。