植物は外界の環境変化に応答することで発生・分化のプログラムを制御しており、特に栄養生長から生殖生長への生長相の移行のタイミングを決定することは植物にとって重要な問題である。この生長相の移行は花成と呼ばれ、特に日長の変化によって花成が誘導される現象は光周性花成誘導と呼ばれている。光周性花成誘導では葉における日長の計測が重要であり、正確な日長の計測には光受容体と概日時計の協調的な働きが必要である。光周性花成誘導における光受容体としては光受容タンパク質であるフィトクロムが主に働いているとされている。しかし、フィトクロムが実際にどのように光周性花成誘導に関与しているかは未だ十分に解明されていない。
フィトクロムについては、赤色光・遠赤色光を吸収し、光可逆性を示すこと等がこれまでの生理学的・分光学的な研究から明らかになってきたものの、分子遺伝学的な知見はあまり得られていなかった。近年、シロイヌナズナをモデル植物とした分子遺伝学的な手法による解析が進み、シロイヌナズナには5種類のフィトクロムが存在し、それぞれが機能分担をしていることが明らかになると同時に、光周性花成誘導における機能等も徐々に明らかになってきている。しかし、シロイヌナズナは条件的長日植物であり、光周期依存的な花成誘導の他にもさまざまな花成誘導経路が複雑に作用し合っている。そこで本研究では、古くから花成制御のモデル植物として用いられてきた絶対的短日植物のアサガオを用いることとした。アサガオは、一回の短日処理で花成を誘導することができ、光周性が非常に強いことが特徴である。本研究では、アサガオを用いてフィトクロムの機能を解析し、光周性花成における光受容体の作用機構の解明を通して光周性花成誘導の分子機構の一端を明らかにすることを目的としている。
【方法】
実験材料として、アサガオ品種「紫」(Pharbitis nil cv. Violet)を用いた。アサガオ品種「紫」の花成は光周性に厳密に制御されており、播種後5日目の芽生えの段階でも1回の短日処理によって花成が誘導できる。また、誘導暗期中に光パルスを与える光中断や誘導暗期直前に遠赤色光(FR)を照射するEOD-FR処理によって花成が著しく阻害されるといった特徴を持っている。さらに、暗黒中で育てた芽生えに光パルスを与えて花成を誘導する骨格光周期も含めたさまざまな光周期条件を用い、アサガオのフィトクロム遺伝子の特性解析を進めた。
1)アサガオ・フィトクロム遺伝子(PnPHY)のシークエンス解析
基礎生物学研究所から分与された全長cDNA及び部分断片のシークエンス解析を行った。部分長のcDNAについては、5’RACE-PCR法を用いて長い断片を増幅し、シークエンスを行った。また、データベースでの相同性検索によって分類を行い、保存ドメインから機能の推定を行った。
2)アサガオ・フィトクロム遺伝子の発現解析
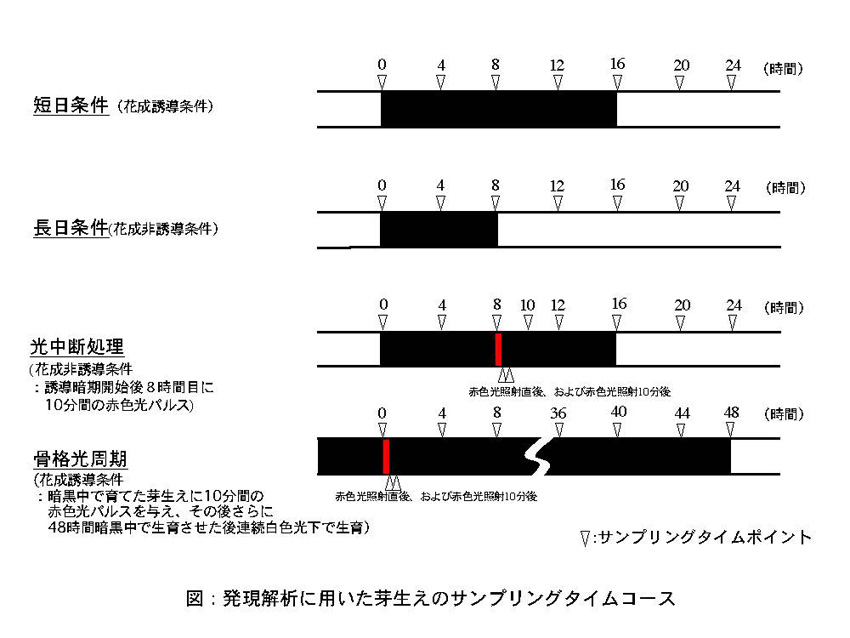
短日処理(花成誘導条件)及び長日処理や光中断処理(花成阻害条件)において発現に変化が見られるフィトクロムを特定するため、Ribonuclease Protection Assay (RPA)による発現解析を行った。
25℃連続光下で生育させた播種後6日目のアサガオに対し、花成誘導条件として16時間の暗期処理を行った個体(短日条件)および非花成誘導条件として8時間の暗期処理を行った個体の各々から4時間おき(長日条件)、誘導暗期4時間おき、誘導暗期開始後8時間目に10分間の赤色光パルスを与えた直後とその20分後(光中断反応後の速い応答)、さらに暗黒中で育てた播種後4日目の芽生えに10分間の赤色光パルスを与え、その後さらに48時間暗黒中で生育させた後連続白色光下で生育させた黄化芽生え(骨格光周期)など、下図に示すようなさまざまな光周期処理条件を与えた個体からtotal RNAを抽出した。このtotal RNAを用いてRPAによるPnPHYの発現解析を行った。
【結果】
1)PnPHYのシークエンス解析
PnPHYEについては既に全長cDNAが単離され、全塩基配列も決定されていたため、その他のPHYA, B, Cについてシークエンス解析を行った。その結果、PnPHYAは1127アミノ酸(a.a.)からなる約125 kDのタンパク質を、また、PnPHYCは1121 a.a.からなる約124 kDのタンパク質をコードしていた。PnPHYBに関しては2種類のcDNAの存在が明らかとなり、それぞれPnPHYB1, およびPnPHYB2と名付けた。PnPHYB1は全長cDNAが単離されていたが、PnPHYB2は5’側の約600bpが欠けていた。この部分断片を5’RACE法を用いて増幅・単離し、シークエンス解析を行った。その結果、PnPHYB1は1131 a.a.からなる約125 kDのタンパク質を、PnPHYB2は1134 a.a.からなる約125 kDのタンパク質をそれぞれコードしていた。
また、データベースを用いた相同性解析の結果、今回解析した5種類のPnPHYは他の植物のフィトクロムと約60?89%の相同性を示し、フィトクロム活性に必要不可欠な発色団であるフィトクロモビリンの結合ドメイン、シグナル伝達関連ドメインとしてタンパク質-タンパク質相互作用に関与するPAS配列、C末端にあるキナーゼ様ドメイン等が保存されていた。これらの機能ドメインの存在から、フィトクロムがフィトクロモビリンと結合して光情報を感受し、その光情報を下流側因子に伝達する光シグナル伝達因子であることが示唆された。
2)PnPHYの発現解析
PnPHYの発現解析に関しては、現在RPA系を立ち上げている段階である。RPAを早急に確立し、短日処理、光中断処理、EOD-FR処理、骨格光周期中における各フィトクロムの発現解析を行い、アサガオの光周性花成誘導に重要な機能を担っているフィトクロムの分子種を同定していきたい。さらに、アサガオ・フィトクロム遺伝子の過剰発現体及びRNAiによる発現抑制体を作製し、フィトクロムと光周性花成誘導との関係を明らかにしていきたい。