|
偮偔偽惗暔僕儍乕僫儖丂Tsukuba Journal of Biology (2005) 4:
TJB200501200100766
揮幨場巕MafG偺DNA擣幆婡峔偺夝柧亅MafG(21-123)/DNA暋崌懱偺X慄寢徎峔憿夝愅
崅愳丂峢柧 乮拀攇戝妛 惗暔妛椶 係擭乯丂丂巜摫嫵堳丗
揷拞丂弐擵 乮拀攇戝妛 惗柦娐嫬壢妛尋媶壢乯
亙彉榑亜
丂Maf孮僞儞僷僋幙偼丄DNA擣幆偵娭傢傞墫婎惈椞堟偲丄擇検壔偵昁梫側儘僀僔儞僕僢僷乕偐傜側傞墫婎惈儘僀僔儞僕僢僷乕乮bZip乯峔憿傪帩偮揮幨挷愡場巕偱偁傞偑丄bZip偺捈慜偵丄EHR(extended homology region)偲屇偽傟傞屌桳偺傾儈僲巁攝楍傪帩偮(恾1)丅Maf孮僞儞僷僋幙偼丄偙偺EHR偲墫婎惈椞堟偵傛偭偰丄MARE偲屇偽傟傞13-14 bp偺DNA攝楍傪摿堎揑偵擣幆偡傞丅MARE偼丄拞墰偵AP1-core(Fos傗Jun側偳偺bZip僞儞僷僋幙偑擣幆偡傞攝楍)傪帩偪丄偦偺椉抂偵3 bp偐傜側傞 flanking 椞堟偑懚嵼偡傞(恾1)丅Maf孮僞儞僷僋幙偼丄懠偺bZip僞儞僷僋幙偲摨條偵墫婎惈椞堟偱AP1-core偲寢崌偡傞偑丄摨帪偵屌桳偺EHR偱flanking椞堟傪擣幆偟丄MARE偲嫮偔寢崌偡傞偲峫偊傜傟偰偄傞丅Maf孮僞儞僷僋幙偺堦偮偱偁傞MafG偺DNA寢崌僪儊僀儞(EHR偲墫婎惈椞堟偐傜側傞)偺峔憿偼丄婛偵摉尋媶幒偱寛掕偟偨偑丄嬶懱揑偵偳偺傛偆側憡屳嶌梡偱MARE傪擣幆偟偰偄傞偐偼枹偩柧傜偐偵側偭偰偄側偄丅偦偙偱丄Maf孮僞儞僷僋幙偺DNA擣幆婡峔傪夝柧偡傞偨傔偵丄EHR偲bZip傪帩偮MafG(21-123)偲丄MARE傪娷傓DNA偲偺暋崌懱偺X慄寢徎峔憿夝愅傪峴偭偨丅
亙曽朄亜
(1) MafG(21-123)偺挷惢
丂摉尋媶幒偱婛偵妋棫偝傟偰偄傞戝挵嬠傪梡偄偨敪尰偲惛惢偺僾儘僩僐乕儖偵廬偭偰丄MafG(21-123)偺戝検敪尰偲惛惢傪峴偭偨丅
(2) MARE傪娷傓擇杮嵔DNA偺挷惢
丂堿僀僆儞岎姺僇儔儉偲扙墫僇儔儉傪梡偄偰惛惢偟偨崌惉僆儕僑僰僋儗僆僠僪 15a(5'-GTGCTGACTCATCAG-3')偲15b(5'-CTGATGAGTCAGCAC-3')傪傾僯乕儕儞僌偟丄擇杮嵔DNA傪摼偨丅
(3) MafG(21-123)/DNA暋崌懱偺寢徎壔
丂MafG(21-123)偲擇杮嵔DNA傪儌儖斾偱2 : 1偲側傞傛偆偵崿崌偟偰挷惢偟偨暋崌懱梟塼(0.2-0.4 mM)傪梡偄丄忲婥奼嶶僴儞僊儞僌僪儘僢僾朄偱寢徎偺嶌惢傪帋傒偨丅
(4) X慄夞愜僨乕僞偺廂廤偲夝愅
丂嶌惢偟偨寢徎偺X慄夞愜僨乕僞偼丄崅僄僱儖僊乕壛懍婍尋媶婡峔(PF-AR, NW12)偵偍偄偰廂廤偟偨丅X慄偺攇挿偼0.978 侌丄専弌婍偼ADSC Quantum 210, 僨乕僞張棟偵偼僾儘僌儔儉HKL2000傪梡偄偨丅
丂
亙寢壥偲峫嶡亜
丂LB攟抧1L偐傜栺25 mg偺惛惢MafG(21-123)偑摼傜傟偨丅偙偺僞儞僷僋幙偲擇杮嵔DNA偐傜側傞暋崌懱偵偮偄偰丄巗斕偺僗僋儕乕僯儞僌僉僢僩傪梡偄偰丄20亷偱寢徎壔傪帋傒偨丅偦偺寢壥丄2-Methyl-2,4-pentanediol偲Polyethylene glycol monomethylether 550傪捑揳嵻偲偡傞忦審偱旝彫寢徎偑摼傜傟偨偺偱丄偙偺忦審傪婎偵寢徎壔忦審傪嵟揔壔偟丄傛傝戝偒偄僒僀僘(栺0.5 mm 亊 0.05 mm 亊 0.1 mm)偺扨寢徎傪摼傞偙偲偵惉岟偟偨(恾2)丅摼傜傟偨榋曽徎偺寢徎傪梡偄偰丄暘夝擻栺4 侌偺X慄夞愜僨乕僞傪廂廤偟偨偲偙傠丄奿巕掕悢偼a = b = 180 侌丄c = 409 侌偱偁偭偨丅尰嵼丄偙偺僨乕僞傪梡偄偰丄MafG偺DNA寢崌僪儊僀儞偺NMR峔憿傪僒乕僠儌僨儖偲偟偨暘巕抲姺朄偵傛傞埵憡寛掕傪帋傒偰偄傞丅傑偨丄傛傝崅暘夝擻偺僨乕僞傪摼傞偨傔偵丄寢徎壔忦審傗寢徎壔偵巊梡偡傞DNA偺挿偝側偳偵偮偄偰専摙拞偱偁傞丅
亙幱帿亜
丂杮尋媶傪峴偆偵偁偨傝丄懡偔偺屼巜摫偲屼嫤椡傪捀偄偨搶杒戝妛 懡尦暔幙壢妛尋媶強 崟壨攷暥攷巑偵怺偔姶幱抳偟傑偡丅
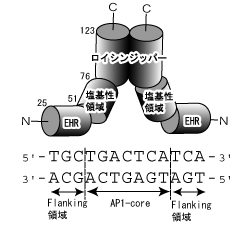
恾1 Maf孮僞儞僷僋幙偺DNA寢崌椞堟偲偦偺擣幆攝楍
丂N偲C偼偦傟偧傟N枛抂偲C枛抂傪丄悢帤偼MafG
丂偵偍偗傞傾儈僲巁巆婎偺斣崋傪帵偡丅

恾2 MafG(21-123)/DNA暋崌懱偺寢徎幨恀
©2005 拀攇戝妛惗暔妛椶
|