|
偮偔偽惗暔僕儍乕僫儖丂Tsukuba Journal of Biology (2005) 4: TJB200501200100787
MafB,c-Maf僟僽儖僲僢僋傾僂僩儅僂僗偺昞尰宆夝愅
挿扟愳丂榓婸乮拀攇戝妛 惗暔妛椶 係擭乯丂丂巜摫嫵堳丗 崅嫶丂抭 乮拀攇戝妛 恖娫憤崌壢妛尋媶壢乯
亂 攚宨丒栚揑 亃
丂戝Maf 孮揮幨場巕偼丄儗僩儘僂僀儖僗AS42偐傜扨棧偝傟偨娻堚揱巕v-maf偺嵶朎娭楢堚揱巕偱偁傝丄儅僂僗偱偼尰嵼傑偱偵c-Maf, MafB, MafA, Nrl偺係庬椶偑摨掕偝傟偰偄傞丅戝Maf孮揮幨場巕偼丄偦偺N枛抂懁偵揮幨妶惈壔擻傪桳偡傞巁惈僪儊僀儞傪帩偪丄偙傟傪帩偨側偄彫Maf孮揮幨場巕偲偼嬫暿偝傟傞丅傑偨C枛抂懁偵偼墫婎惈僪儊僀儞偲儘僀僔儞僕僢僷乕僪儊僀儞偐傜側傞bZIP僪儊僀儞傪帩偭偰偍傝丄偙偺bZIP僪儊僀儞傪夘偟偰儂儌僟僀儅乕傗僿僥儘僟僀儅乕傪宍惉偟丄Maf擣幆攝楍傊偲寢崌偡傞偙偲偵傛偭偰堚揱巕敪尰傪惂屼偟偰偄傞丅摉尋媶幒偱偼c-Maf丄媦傃MafB寚懝儅僂僗傪偦傟偧傟嶌惢丄夝愅偟偰偍傝丄c-Maf寚懝儅僂僗偼娽偺敪惗偵偍偗傞悈徎懱慇堐嵶朎偺暘壔傗丄T嵶朎偵傛傞IL-4偺嶻惗側偳偵堎忢偑尒傜傟傞丅傑偨MafB寚懝儅僂僗偼恡巺媴懱懌嵶朎偵偍偗傞懌撍婲偺寚擛傗暯峵姶妎傪巌傞撪帹偺宍惉晄慡丄偝傜偵墑悜屇媧拞悤僯儏乕儘儞偺敪惗堎忢側偳偑尒傜傟傞丅偙偺俀庬椶偺堚揱巕寚懝儅僂僗偼丄嫟偵惗屻捈屻偱巰朣偟偰偟傑偆丅
丂彫Maf孮揮幨場巕傪巒傔偄偔偮偐偺堚揱巕孮偱偼丄僼傽儈儕乕堚揱巕傪暋悢屄寚懝偝偣傞偙偲偵傛偭偰扨撈寚懝儅僂僗偱偼偁傜傢傟側偐偭偨昞尰宆偑尠嵼壔偡傞偙偲偑傢偐偭偰偄傞丅偙傟傑偱偺尋媶偐傜丄彮側偔偲傕儅僋儘僼傽乕僕偱偼MafB偲c-Maf偑嫟偵敪尰偟偰偄傞偙偲偑敾柧偟偰偄偨偑丄MafB偲c-Maf偺敪惗夁掱偵偍偗傞徻嵶側敪尰僷僞乕儞偺夝愅偑惉偝傟偰偄側偄慻怐傕懡偔丄MafB偲c-Maf傪嫟偵寚懝偝偣傞偙偲偵傛偭偰昞尰宆偑廳撃壔偟偨傝丄怴偨側堎忢偑惗偠傞壜擻惈偑偁偭偨丅廬偭偰杮尋媶偱偼MafB-/-::c-Maf-/-儅僂僗傪夝愅偡傞偙偲偵傛傝丄偦傟偧傟偺扨撈寚懝儅僂僗偱偼傢偐傜側偐偭偨in vivo偱偺MafB偲c-Maf偺婡擻傪夝柧偡傞偙偲傪栚揑偲偡傞丅
亂曽朄亃
MafB+/-::c-Maf+/-儅僂僗摨巑傪岎攝偟丄戀惗10.5擔乣戀惗12.5擔偺MafB-/-::c-Maf-/-戀帣傪摼偨丅偙傟傪嵽椏偲偟偰埲壓偺傛偆側幚尡傪峴偭偨丅
丂丒HE愼怓
戀帣偺僷儔僼傿儞愗曅傪嶌惢偟偨偁偲丄僿儅僩僉僔儕儞偲僄僆僔儞偱嵶朎傪愼怓偟偰慻怐偺娤嶡傪峴偭偨丅
丂丒TUNEL愼怓
戀帣偺僷儔僼傿儞愗曅傪嶌惢偟丄FITC偱儔儀儖偟偨dUTP傪僞乕儈僫儖僨僆僉僔僰僋儗僆僠僕儖僩儔儞僗僼僃儔乕僛傪梡偄偰DNA抐曅3'懁偵晅壛偟偨丅
偝傜偵HRP偺寢崌偟偨anti-FITC峈懱傪嶌梡偝偣偨屻丄HRP偺婎幙偱偁傞DAB偱敪怓偝偣偰傾億僩乕僔僗嵶朎傪壜帇壔偟偨丅
丂丒僐儘僯乕傾僢僙僀
戀惗10.5擔偺棏墿擷偲戀惗12.5擔偺娞嵶朎傪丄憹怋傪懀偡偄偔偮偐偺僒僀僩僇僀儞傪壛偊偨儊僠儖僙儖儘乕僗攟抧偵傑偄偰丄惉挿偟偰偒偨愒寣媴宯偺僐儘僯乕乮
CFU-E乯悢傪寁應偟偨丅
亂寢壥丒峫嶡亃
丂MafB+/-::c-Maf+/-儅僂僗摨巑傪岎攝偟偰c-Maf-/-::MafB-/-儅僂僗傪嶌惢偟偨偲偙傠丄戀惗12.5擔偱偼昻寣偺徢忬傪帵偟乮恾侾乯丄偦傟偧傟偺扨撈寚懝儅僂僗傛傝傕憗婜偺戀惗拞婜偱抳巰偲側傞偙偲偑傢偐偭偰偒偨丅師偵丄昻寣偺尨場傪扵傞偨傔慻怐妛揑側夝愅傪峴偭偨偲偙傠丄擇師憿寣偺応偱偁傞娞憻偱愒寣媴宯偺嵶朎偑傾億僩乕僔僗傪婲偙偟偰偄傞偙偲偑柧傜偐偲側偭偨丅戀惗12.5擔偺娞嵶朎偱僐儘僯乕傾僢僙僀傪峴偭偨偲偙傠丄栰惗宆偵斾傋偰娞嵶朎悢偁偨傝偺CFU-E偺悢偑憹壛偟偰偄偨丅偟偐偟側偑傜丄戀惗10.5擔偺棏墿擷偱僐儘僯乕傾僢僙僀傪峴偭偨応崌偼栰惗宆偲偺堘偄偼尒傜傟側偐偭偨丅偙傟傜偺寢壥偐傜丄MafB-/-::c-Maf-/-儅僂僗偱尒傜傟偨愒寣媴宯嵶朎偺傾億僩乕僔僗偼丄愒寣媴帺懱偵尨場偑偁傞偺偱偼側偔丄戀帣娞憻偵偍偗傞旝彫娐嫬偺攋抅偑尨場偱偁傞偲峫偊傜傟傞丅崱屻偼丄MafB-/-::c-Maf-/-儅僂僗偺娞憻偵偍偄偰傾億僩乕僔僗偑婲偙傞徻嵶側暘巕儊僇僯僘儉傪夝愅偟偰偄偔梊掕偱偁傞丅
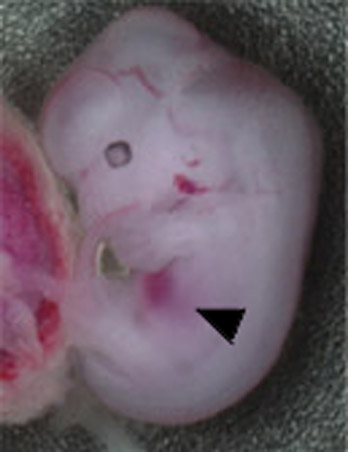 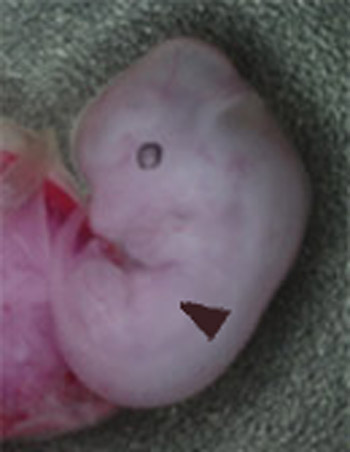 恾侾丂戀惗12.5擔丂MafB+/-::c-Maf+/-乮嵍乯偲
恾侾丂戀惗12.5擔丂MafB+/-::c-Maf+/-乮嵍乯偲
丂丂丂MafB-/-::c-Maf-/-乮塃乯偺戀帣丅
僐儞僩儘乕儖乮嵍乯偵斾傋偰丄僟僽儖僲僢僋傾僂僩儅僂僗乮塃乯偼娞憻乮栴摢乯偑敀偔丄恎懱偺戝偒偝傕彫偝偄丅
©2005 拀攇戝妛惗暔妛椶
|