|
偮偔偽惗暔僕儍乕僫儖丂Tsukuba Journal of Biology (2006) 5: TJB200601200200763
僶僜僾儗僔儞偵傛傞僆儗僉僔儞恄宱妶惈壔儊僇僯僘儉偺揹婥惗棟妛揑夝愅
忢徏丂桭旤乮拀攇戝妛 惗暔妛椶 係擭乯丂丂巜摫嫵堳丗嶳拞丂復峅乮拀攇戝妛 恖娫憤崌壢妛尋媶壢乯
尋媶偺攚宨
丂僆儗僉僔儞偼1998擭偵G僞儞僷僋幙嫟栶宆庴梕懱偺撪場惈儕僈儞僪偲偟偰摨掕偝傟偨恄宱儁僾僠僪偱偁傝丄悋柊丒妎惲偺挷愡傗愛怘峴摦偺惂屼側偳偺廳梫側惗棟婡擻傪扴偭偰偄傞丅僆儗僉僔儞嶻惗恄宱偼帇彴壓晹奜懁栰偵摿堎揑偵嬊嵼偟丄偦偺搳幩愭偼擼偺峀斖偵傢偨偭偰偄傞丅堦曽偱丄僆儗僉僔儞恄宱偺妶摦傪挷愡偡傞媮怱惈偺儊僇僯僘儉偼枹偩廫暘夝柧偝傟偰偄側偄丅杮尋媶偱偼丄僆儗僉僔儞恄宱摿堎揑偵Enhanced Green Fluorescent Protein (EGFP)傪敪尰偡傞僩儔儞僗僕僃僯僢僋儅僂僗乮僆儗僉僔儞/EGFP儅僂僗乯傪梡偄偰丄僆儗僉僔儞恄宱偵懳偡傞僶僜僾儗僔儞偺嶌梡傪僗儔僀僗僷僢僠僋儔儞僾朄偵傛偭偰徻嵶偵夝愅偡傞偙偲傪帋傒偨丅
嵽椏偲曽朄
丂2-4廡椷偺僆儗僉僔儞/EGFP儅僂僗傪梡偄偰幚尡傪峴偭偨丅廫暘偵杻悓偟偨儅僂僗偐傜偡偽傗偔擼傪揈弌偟丄昘椻偟偨僗僋儘乕僗儀乕僗偺Hepes-Buffer拞偱岤偝350兪m偺僗儔僀僗昗杮傪價僽儔僩乕儉傪梡偄偰嶌惢偟偨丅摼傜傟偨僗儔僀僗偼墫壔僫僩儕僂儉儀乕僗偺Hepes-Buffer拞偱1帪娫僀儞僉儏儀乕僩偟偨屻丄幚尡偵梡偄偨丅EGFP傪敪尰偟偨僆儗僉僔儞恄宱偼丄寀岝尠旝嬀壓偱摨掕偟丄嬤愒奜旝暘姳徛尠旝嬀憸傪尒側偑傜丄儂乕儖僙儖僷僢僠僋儔儞僾朄偵傛傝偦偺妶摦傪婰榐偟偨丅栻塼偼嬊強搳梌朄偵傛傝丄僆儗僉僔儞恄宱偺嬤朤偐傜搳梌偟偨丅
寢壥偲峫嶡
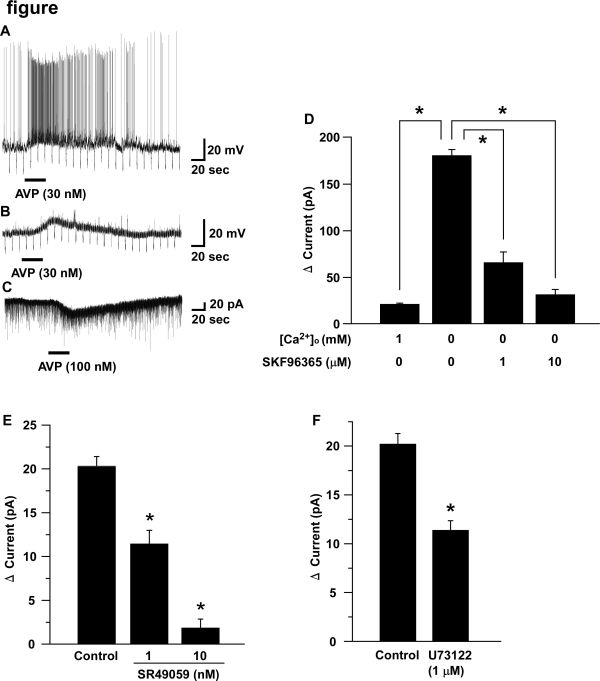
丂僇儗儞僩僋儔儞僾偵傛偭偰僆儗僉僔儞恄宱偺枌揹埵傪儌僯僞乕偟側偑傜丄僶僜僾儗僔儞乮30 nM乯傪搳梌偡傞偲丄扙暘嬌偲偦傟偵敽偆敪壩昿搙偺憹壛偑尒傜傟偨乮fig. A乯丅扙暘嬌偼僥僩儘僪僩僉僔儞懚嵼壓偱傕娤嶡偝傟乮fig. B乯丄偦偺斀墳偼擹搙埶懚揑偱偁偭偨偙偲偐傜捈愙嶌梡偲峫偊傜傟偨丅傑偨丄儃儖僥乕僕僋儔儞僾偵傛偭偰枌揹埵傪-60 mV偵屌掕偟偰僶僜僾儗僔儞(100 nM)傪搳梌偡傞偲丄20 pA慜屻偺撪岦偒揹棳偑娤嶡偝傟偨乮fig. C乯丅僶僜僾儗僔儞偵傛偭偰桿敪偝傟傞撪岦偒揹棳傪徻嵶偵夝柧偡傞栚揑偱丄斀揮揹埵傪媮傔偨偲偙傠栺0 mV嬤曈偱偁偭偨丅偝傜偵丄嵶朎奜偺僇儖僔僂儉傪彍嫀偡傞偲僶僜僾儗僔儞偵傛傞撪岦偒揹棳偑憹戝偟丄旕慖戰惈梲僀僆儞僠儍僱儖偺慾奞嵻偱偁傞SKF96365偵傛偭偰丄偙偺撪岦偒揹棳偼擹搙埶懚揑偵梷惂偝傟偨乮fig. D乯丅偟偨偑偭偰丄僶僜僾儗僔儞偵傛偭偰桿敪偝傟傞撪岦偒揹棳偼丄旕慖戰惈梲僀僆儞僠儍僱儖偑奐岥偡傞偙偲偱惗偠偰偄傞偲峫偊傜傟傞丅
丂師偵僆儗僉僔儞恄宱偺妶惈壔偵娭梌偟偰偄傞僶僜僾儗僔儞庴梕懱傪栻棟妛揑偵摨掕偟偨丅僶僜僾儗僔儞庴梕懱偲偟偰3庬椶偺僒僽僞僀僾乮V1a丒V1b丒V2乯偑抦傜傟偰偄傞丅V1a庴梕懱偺慖戰揑漢峈栻偱偁傞SR49059偑丄擹搙埶懚揑偵僶僜僾儗僔儞偵傛傞撪岦偒揹棳傪梷惂偟偨乮fig. E乯丅V1a庴梕懱偼G僞儞僷僋幙嫟栶宆庴梕懱偱偁傝丄嶰検懱G僞儞僷僋幙偺偆偪G兛q僒僽僋儔僗偲嫟栶偟偰偄傞偙偲偑曬崘偝傟偰偄傞丅偦偙偱丄嵶朎撪僔僌僫儖揱払宱楬傪柧傜偐偵偡傞偨傔丄儂僗儂儕僷乕僛C偺慾奞嵻偱偁傞U73122傪嶌梡偝偣偨丅U73122偼僶僜僾儗僔儞偵傛傞撪岦偒揹棳傪梷惂偟偨偙偲偐傜乮fig. F乯丄庴梕懱偐傜旕慖戰惈梲僀僆儞僠儍僱儖傑偱偺僔僌僫儖揱払宱楬偵儂僗儂儕僷乕僛C偺妶惈壔偑娭梌偟偰偄傞偲峫偊傜傟傞丅
丂杮尋媶偵傛傝丄僶僜僾儗僔儞偑V1a庴梕懱傪夘偟偨旕慖戰惈梲僀僆儞僠儍僱儖偺奐岥偵傛偭偰丄僆儗僉僔儞恄宱傪扙暘嬌偝偣偰偄傞偙偲偑柧傜偐偲側偭偨丅崱屻偼偙偺僶僜僾儗僔儞恄宱偵傛傞僆儗僉僔儞恄宱傊偺嫽暠惈擖椡偺惗棟揑堄媊偺夝柧傪峴偆梊掕偱偁傞丅
©2006 拀攇戝妛惗暔妛椶
|