|
偮偔偽惗暔僕儍乕僫儖丂Tsukuba Journal of Biology (2006) 5: TJB200601200200767
揮幨場巕Runx3偺枛徑恄宱敪惗偵偍偗傞婡擻夝愅
惣懞丂旤崄 乮拀攇戝妛 惗暔妛椶 係擭乯丂丂巜摫嫵堳丗巙夑丂棽 乮拀攇戝妛 恖娫憤崌壢妛尋媶壢乯
亂栚揑偲攚宨亃
丂RUNX僼傽儈儕乕揮幨場巕偼丄僔儑僂僕儑僂僶僄segmentation gene偺runt偲儂儌儘僕乕偺崅偄Runt僪儊僀儞傪帩偮揮幨場巕偱偁傝丄歁擕椶偱偼Runx1乣3偺俁偮偑摨掕偝傟偰偄傞丅嬤擭偺尋媶偐傜丄Runx1偼庡偵憿寣姴嵶朎偵敪尰偟丄媫惈崪悜惈敀寣昦偺尨場堚揱巕偺侾偮偲偟偰丄堦曽Runx2偼崪夎嵶朎偵敪尰偟丄嵔崪摢奧堎宍惉徢偲偺娭楢偵偍偄偰丄傑偨Runx3偼徚壔娗擲枌忋旂嵶朎偵敪尰偟丄堓僈儞偺梷惂堚揱巕偲偟偰丄堚揱巕偺堎忢偲幘姵偲偺娭楢偑曬崘偝傟偰偍傝丄Runx僼傽儈儕乕揮幨場巕偼丄嵶朎偺敪惗丒暘壔夁掱偵偍偄偰嬌傔偰廳梫側栶妱傪帩偮偲峫偊傜傟偰偄傞丅
丂偲偙傠偱丄Runx1偲Runx3偼敪惗夁掱偺堦晹偺恄宱宯偵偍偄偰傕敪尰偡傞偙偲偑傢偐偭偰偄傞偑丄偦偺婡擻偵偮偄偰偼枹偩晄柧側揰偑懡偄丅偦偙偱摉尋媶幒偱偼丄Runx3堚揱巕寚懝儅僂僗偺宍懺妛揑夝愅傪峴側偄丄偙偺儅僂僗偱偼屌桳姶妎惈DRG僯儏乕儘儞偺幉嶕偑拞悤懁偺昗揑偱偁傞愐悜慜妏偲枛徑懁偺昗揑偱偁傞嬝朼悗傑偱摓払偣偢偵搑拞偱徚幐偟丄恄宱夞楬宍惉偵堎忢偑偁傞偙偲傪柧傜偐偵偟偨乮Inoue, Ozaki, Shiga et al. 2002乯丅側偍DRG僯儏乕儘儞悢偵尠挊側曄壔偼尒傜傟側偐偭偨丅堦曽丄Levanon傜(2002)偼丄暿屄偵嶌弌偝傟偨摨堚揱巕寚懝儅僂僗偺夝愅偐傜丄屌桳姶妎惈DRG僯儏乕儘儞偺暘壔偲惗懚偺忈奞偵傛傞僯儏乕儘儞悢偺尭彮傪曬崘偟偰偍傝丄摉尋媶幒偺寢壥偲偺堘偄偺棟桼偼晄柧偱偁傞丅
丂偦偙偱杮尋媶偱偼丄屌桳姶妎惈DRG僯儏乕儘儞偺儅乕僇乕偱偁傞parvalbumin乮PV乯偵拲栚偟丄柶塽慻怐壔妛愼怓朄傪梡偄偰丄Runx3堚揱巕寚懝儅僂僗偺夝愅傪峴偄丄偙偺僯儏乕儘儞偺敪惗丒暘壔偵偍偗傞Runx3偺栶妱傪夝柧偡傞偙偲傪栚揑偲偡傞丅
亂曽朄亃
丂Runx3堚揱巕寚懝儅僂僗偼僕乕儞僞乕僎僢僥傿儞僌朄偵傛傝婛偵嶌弌偝傟偰偄傞傕偺傪梡偄偨丅Runx3-/-儅僂僗偼庼擕偑偱偒偢丄惗屻傑傕側偔巰朣偡傞丅偦偙偱Runx3+/-儅僂僗摨巑傪岎攝偟丄惗屻0擔栚偺屄懱傪僒儞僾儕儞僌偟偨丅側偍埲壓丄Runx3+/+儅僂僗丄Runx3-/-儅僂僗偼丄摨堦忦審壓偵偍偄偰偄傞丅
丂係亾僷儔僼僅儖儉傾儖僨僸僪/儕儞巁娚徴塼傪屌掕塼偲偟丄怱寣塼弞娐宯傪棙梡偡傞燇棳屌掕傪峴偄丄2斢屻屌掕傪峴偭偨丅戞12榏崪傪巜昗偵丄嫻晹DRG傪戞2嫻悜(Th2)偐傜Th侾俀偵傢偨偭偰3愡偛偲偵愗傝弌偟丄扙悈摟揙偺屻丄僷儔僼傿儞偵曪杽偟偨丅偦偺屻丄儈僋儘僩乕儉偱8兪m偺楢懕愗曅傪嶌惉偟偨丅奺僒儞僾儖偺DRG儗儀儖傪専嬀偵偰妋擣偟丄摨堦儗儀儖偺DRG傪夝愅偵梡偄偨丅扙僷儔僼傿儞偺屻丄0.3%夁巁壔悈慺悈/儊僞僲乕儖偱撪嵼惈儁儖僆僉僔僞乕僛妶惈傪彍嫀偟丄懕偄偰5亾惓忢儎僊寣惔/儕儞巁娚徴塼偱擇師峈懱偺旕摿堎揑寢崌傪攔彍偟偨丅堦師峈懱偲偟偰丄Rabbit 峈PV峈懱乮1: 500, Swant乯傪斀墳偝偣丄懕偄偰擇師峈懱偲偟偰丄價僆僠儞昗幆峈Rabbit IgG峈懱乮1: 300, Vector乯傪斀墳偝偣偨丅偝傜偵ABC朄偵傛傝僔僌僫儖傪憹嫮偝偣丄惣梞儚僒價儁儖僆僉僔僞乕僛妶惈偵傛傝DAB敪怓傪峴偭偨丅晻擖屻丄枩擻尠旝嬀乮Axio plan2 imaging, Carl Zeiss幮惢乯偱幨恀傪嶣塭偟丄DRG偵偍偄偰愼怓偝傟偨僯儏乕儘儞傪慡偰寁應偟偨丅偙偺偲偒丄暋悢偺愗曅偵傑偨偑傞傕偺偼丄侾屄偲偟偰庢傝埖偭偨丅
丂
亂寢壥偲峫嶡亃
丂惗屻0擔栚偺Th11偵偍偄偰栰惗宆乮3屄懱丄6DRG乯偱偼丄暯嬒144.7亇15.3屄乮p< 0.001乯偺PV梲惈僯儏乕儘儞偑妋擣偝傟偨丅堦曽丄Runx3寚懝宆乮摨忋乯偱偼擣傔傜傟側偐偭偨丅傑偨丄栰惗宆偱偼丄PV梲惈僯儏乕儘儞偺幉嶕偼丄愐悜偺屻嶕偐傜慜妏傊搳幩偟偰偄偨偑丄Runx3寚懝宆偱偼擣傔傜傟側偐偭偨丅
丂埲忋偺寢壥傛傝丄揮幨場巕Runx3偼丄PV偺敪尰挷愡傪捠偟偰丄屌桳姶妎惈DRG僯儏乕儘儞偺敪惗丒暘壔偵婡擻偡傞壜擻惈偲丄偙偺僯儏乕儘儞偺惗懚傗幉嶕搳幩偵婡擻偡傞壜擻惈偺俀偮偑峫偊傜傟傞丅
丂傑偨丄栰惗宆丄Runx3寚懝宆偲傕偵嬝慄堐偵PV偺敪尰偑擣傔傜傟偨丅偙偺偙偲偐傜丄Runx3寚懝宆偺DRG偵偍偄偰PV梲惈僯儏乕儘儞偑擣傔傜傟側偄偙偲偑丄屌掕忦審側偳偵傛傞愼怓偺栤戣偱偁傞壜擻惈傪攔彍偱偒傞丅偝傜偵丄尦棃Runx3堚揱巕傪敪尰偟側偄嬝慄堐偲偺PV偺敪尰婡彉偺斾妑偐傜丄DRG僯儏乕儘儞偱偼丄Runx3偼傛傝捈愙揑偵PV偺敪尰偵娭梌偟偰偄傞偲峫偊傜傟傞丅
亂崱屻偺曽恓亃
丂Runx3堚揱巕偼戀惗11.5擔栚傛傝敪尰偡傞偙偲偑曬崘偝傟偰偄傞偙偲偐傜丄傑偢丄戀惗15.5擔偵偍偄偰摨條偺夝愅傪峴偄丄忋偺俀偮偺壜擻惈偵偮偄偰扵傞丅
丂Runx3堚揱巕偺敪尰偑尒傜傟傞偄偢傟偺敪払抜奒偵偍偄偰傕丄摨堚揱巕寚懝宆偱PV梲惈僯儏乕儘儞偑擣傔傜傟側偗傟偽丄揮幨場巕Runx3偼屌桳姶妎惈DRG僯儏乕儘儞偵偍偄偰丄PV偺敪尰偲偙偺DRG僯儏乕儘儞偺暘壔偵婡擻偟偰偄傞偲峫偊傜傟傞丅堦曽丄PV梲惈僯儏乕儘儞偑擣傔傜傟傞敪惗抜奒偑偁傞側傜偽丄揮幨場巕Runx3偼偙偺DRG僯儏乕儘儞偺惗懚偲幉嶕搳幩偵婡擻偟偰偍傝丄暘壔偵偼娭梌偟側偄偲峫偊傜傟傞丅
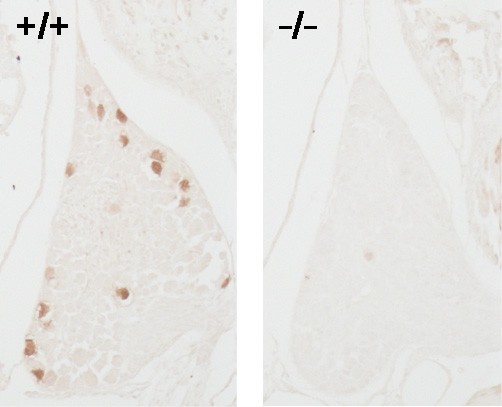
恾侾: 惗屻0擔栚丂Runx3+/+媦傃Runx3-/-儅僂僗偵偍偗傞
parvalbumin梲惈愐悜恄宱愡僯儏乕儘儞悢偺曄壔
©2006 拀攇戝妛惗暔妛椶
|