|
つくば生物ジャーナル Tsukuba Journal of Biology (2006) 5: TJB200601200200786
カタユウレイボヤ精子における補体系C6様タンパク質の解析
水野 克俊 (筑波大学 生物学類 4年) 指導教員:稲葉 一男 (筑波大学 生命環境科学研究科)
【背景と目的】
補体系は、高等脊椎動物において感染初期の生体防御に重要な役割を果たしている自然免疫システムである。近年、補体分子がウニやホヤなどの無脊椎動物にも存在することが明らかになり、その進化系統学的意義が注目されている。一方、さまざまな補体制御因子が精子表面に存在していることが哺乳類で知られており、補体系と受精の関連が指摘されている。体内受精時に雌の免疫系による精子への攻撃を回避する働きがあると考えられているが、真の生理学的意義や分子機能はほとんど明らかにされていない。
尾索動物であるホヤには、獲得免疫系は存在しないが自然免疫系は存在し、さまざまな補体因子を有している。脊椎動物と同じ脊索動物に属している点や、受精において自家不和合性が存在する点などから、ホヤの補体系は免疫系の進化という観点で重要である。当研究室における以前の研究により、カタユウレイボヤの精子細胞膜に、補体系因子の一つであるC6と相同性を示す分子が存在することが示唆された。この分子の研究により、体外受精における補体系の関与について何らかの知見が得られることが期待されると同時に、免疫系と生殖系の進化系統学的な関連性を知る上でも重要であると考えられる。
本研究では、受精における補体系の役割を明らかにする目的で、カタユウレイボヤ精子に存在することが示唆されている補体系C6様タンパク質に対する抗体を作製した。さらに、ウエスタンブロッティングと間接蛍光抗体法により、精子におけるC6様タンパク質の局在を調べた。
【方法】
ホヤのcDNAクラスター10410に属するクローン(ciad016m22)から、PCRによりタンパク質コード領域を増幅し、pET32aベクターにサブクローニングした。これを、エレクトロポレーションにより大腸菌AD494株に導入し、IPTG添加によりチオレドキシン融合タンパク質を発現させた。大腸菌から発現タンパク質を可溶化した後、Hisタグを利用して融合タンパク質のアフィニティ精製を行った。精製した融合タンパク質を等量のアジュバンドと混合し、マウスの皮下に10日間隔で4回注射することにより、C6様タンパク質に対する抗体を作製した。得られた抗体を用いて、カタユウレイボヤ精子に対するウエスタンブロッティングを行った。また、間接蛍光抗体法により、精子におけるC6様タンパク質の局在を調べた。
【結果と考察】
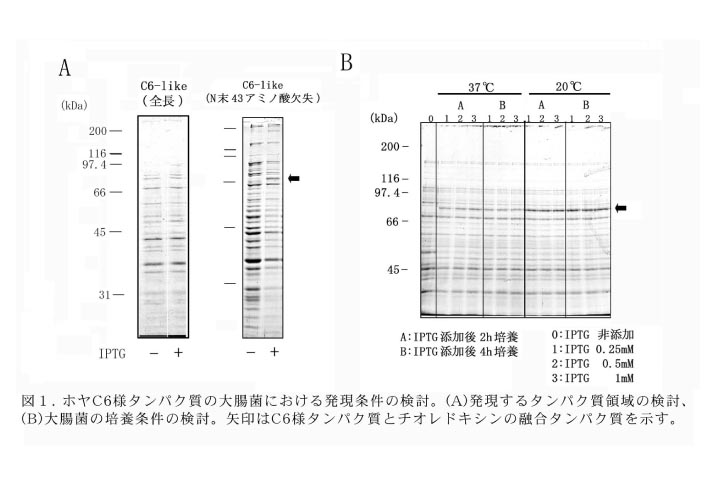
(1)C6様タンパク質の大腸菌における発現(図1)
ciad016m22がコードするタンパク質の全長に対して、タンパク質の発現を試みたところ、IPTG誘導後、顕著なタンパク質の発現は見られなかった。そこで、膜貫通領域とアルギニン/プロリンに富む特徴的な配列を含むN末端の43アミノ酸を含まない領域の発現を試みたところ、予想される分子質量75kDaとほぼ一致するサイズのタンパク質の発現誘導が見られた。発現量が少なかったため、大腸菌の温度、培養時間、およびIPTGの濃度について検討した結果、20℃での培養が発現量の増加に効果的であることがわかった。
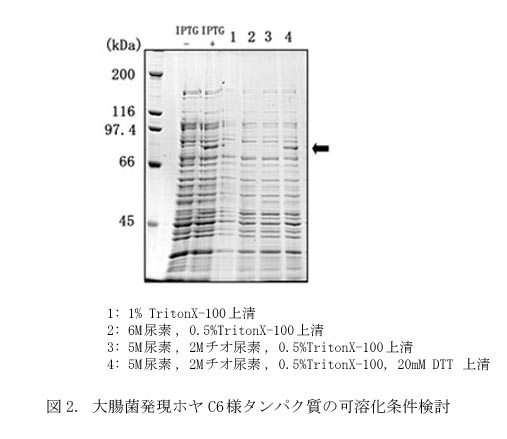
(2)発現タンパク質の可溶化(図2)
大腸菌を回収し、C6様タンパク質とチオレドキシンの融合タンパク質の抽出を試みたところ、6 M 尿素を含んだ溶液でも十分に可溶化されないことがわかった。そこで、尿素より強力な変性剤であるチオ尿素とSH剤であるDTT(ジチオスレイトール)を添加したところ、DTTの存在により効率的に可溶化することができた。
(3)抗体の作製
実験開始当初は、十分量の抗原量が得られず、抗原の精製度も低かった。このような抗原をマウスに免疫して抗体を得たが、カタユウレイボヤ精子に含まれる総タンパク質に対してウエスタンブロッティングを行ったところ、複数のタンパク質が認識されることがわかった。間接蛍光抗体法では精子全体にシグナルが見られたが、これが実際にC6様タンパク質の局在を示しているのであれば興味深い結果である。現在、上記(1), (2)の条件検討に基づき、精製度の高い十分量の抗原を調製し、抗体の作製を再度試みている。得られた抗体を用いて、ウエスタンブロッティングと間接蛍光抗体法を追試する予定である。
【今後の課題】
抗体による受精阻害実験、精子内でC6様タンパク質と相互作用するタンパク質の解析、卵側の結合分子の検索などを行うことにより、受精における精子C6様分子の役割を明らかにする。
©2006 筑波大学生物学類
|