|
つくば生物ジャーナル Tsukuba Journal of Biology (2006) 5: TJB200603SE1.
動物型から植物型へ1世代で変身
―植物進化の謎に迫る不思議な生物「ハテナ」―
TJB学生編集部 (筑波大学 生物学類)
昨年、本学大学院生命環境科学研究科長の井上勲教授とその研究グループの一員であった岡本典子さん(現在オーストラリア、メルボルン大学の研究室に所属)による新種の生物についての論文が米国科学誌「サイエンス」10月14日号に掲載され話題となった。その生物の名は「ハテナ」。
ハテナは長さ約30μmの単細胞生物で、鞭毛(べんもう)を使って動き回る「鞭毛虫」である。ハテナの注目すべき特徴は、細胞内に緑色の藻類が共生して光合成を行っており、分裂して増殖する際に、この共生体が一方の細胞にだけ受け継がれる点である。分裂してできた2つの娘(じょう)細胞の一方は藻類として光合成を行い(植物的生活)、もう一方は捕食性の原生生物として生活する(動物的生活)。このような二重の生活環を持つ生活様式は、植物が多様化する過程の初期段階と考えられ、その点で「ハテナ」は植物進化上の重要な鍵を握る生物とされるのである。
植物の多様性は「二次共生」で生まれた。
植物は、葉緑体により酸素発生型光合成を行う生物群で、陸上植物と多様な藻類のグループが含まれる。実は、葉緑体は元々独立した生物で、その起源は酸素発生型光合成をする唯一の原核生物(核を持たない生物)、シアノバクテリア(藍藻)である。太古の地球で、無色の真核生物(核を持つ生物)がシアノバクテリアを取り込み、葉緑体とすることで「一次植物」と呼ばれる最初の植物群が生まれた。この細胞共生を「一次共生」とよぶ。
植物は細胞の核のDNA以外に、葉緑体にもDNA(取り込まれたシアノバクテリア由来)をもち、これが一次共生を裏付ける証拠ともいえる。現生植物の葉緑体DNAを解析した結果、植物は共通の祖先を持つことがわかっている。
しかしここで矛盾が生じる。植物の核DNAを解析し系統分類すると、植物には様々な起源があることになるのだ。つまり葉緑体でみると単系統なのに、核でみると多系統なのである。この矛盾を解決するのが「二次共生」という考えである。
二次共生とは、一次共生によって誕生した一次植物を、無色の真核生物が取り込むことである。共生体となった一次植物の核は退化し、DNAは残すが核としての機能を失った「ヌクレオモルフ」と呼ばれる痕跡となり、やがて進化と共に消失する。クリプト植物(クリプト藻)のようにヌクレオモルフが残っている植物もあり、二次共生が起きた証拠となっている。
二次共生で誕生した「二次植物」の核は、宿主である無色の真核生物の核である。いろいろな生物が宿主となったため、現生植物の核には様々な系統がみられるのである。つまり現在の植物の多様性は、二次共生によって生まれたといえる。
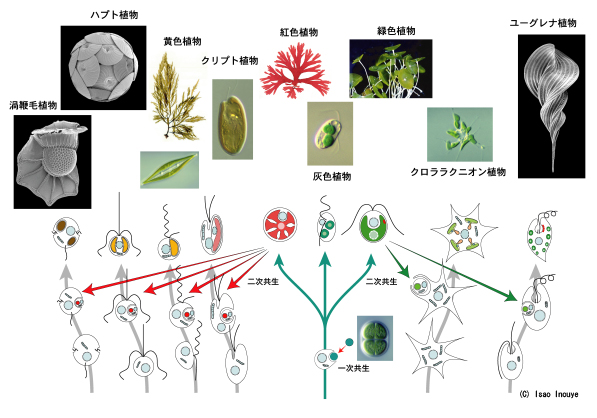
【図1】一次共生・二次共生の流れ
一次共生では無色の真核生物にシアノバクテリアが共生して一次植物が誕生した。 一次植物は単一系統である。一次植物がさまざまな系統の無色真核生物に共生して二次植物が誕生し、写真の植物群のような多様性が生じた。©井上勲
ハテナには藻類が「二次共生」していた。
発見されたハテナは細胞が緑色で、一見、緑色藻類のようだが、2本の鞭毛を使ってシャクトリムシのような動きをしていた。緑色藻類にこのような動きをするものはない。井上教授らがハテナを電子顕微鏡で観察したところ、緑色の部分は、核を持った真核藻類が、鞭毛虫の細胞内に共生しているものとわかった。さらに分裂中の個体の観察では、分裂後の一方の細胞にだけこの藻類が受け継がれていた。通常の藻類では、細胞分裂の前にまず葉緑体が分裂し、分裂後それぞれの娘細胞に葉緑体が分配される。
ハテナの細胞構造が、カタブレファリスという鞭毛虫に似ていることから、カタブレファリスの核DNAを解析したところ、これはほかのどの真核生物とも違うことが判明し、新たな分類門として「カタブレファリス門」が確立された。そしてハテナの核DNA解析から、ハテナはこのカタブレファリス門に属することが確認された。
一方、共生体である緑色藻類は、DNA解析の結果、緑色植物の最も原始的なグループであるプラシノ藻のネフロセルミス属の一種であることがわかった。こうした解析によって、ハテナは無色の真核生物(鞭毛虫)が真核の緑色藻類(プラシノ藻)を共生体として取り込んだ「二次植物」であるということが示されたのである。
分裂後に捕食型生物となったハテナの餌、すなわち共生体となるべきネフロセルミス属の種類が特定・単離収集できていないため、ハテナの培養はいまだに成功していない。
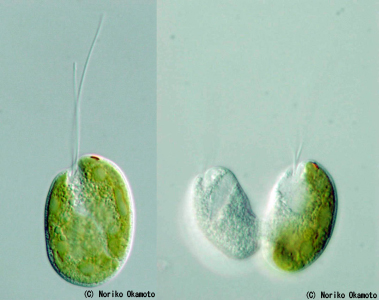 
左【図2】ハテナの光学顕微鏡写真
自然界で見つかるハテナの多くが、この写真 の個体のように、共生体として緑色藻類を細胞内に取り込んでいる。右上の赤い点が眼点。共生藻類を持たない無色のハテナでは、眼点の代わりに捕食装置を持つ。©岡本典子
中【図3】ハテナ分裂終期の写真
ハテナでは宿主と共生体の分裂が同調しておらず、共生体は分裂後、片方の娘細胞(右側)にだけ受け継がれる。もう片方の娘細胞(左側)は共生藻類を持たない無色の個体となる。©岡本典子
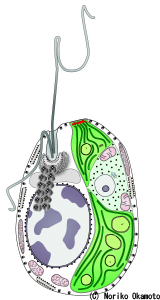
右【図4】ハテナの内部構造(共生藻類を持つ個体)
細胞内左側の大きな円形の構造が核、右側の緑色の部分が共生する緑色藻類である。共生藻類の内部の中央やや上寄りの円形構造が、共生藻類の核である。©岡本典子
動物型のハテナが捕食により植物型に。
ハテナが二次植物の構造を持つことはわかったが、最も重要な謎が残っている。それはハテナの分裂の仕方である。普通の二次植物は、葉緑体分裂後に核分裂が起き、分裂後の2つの娘細胞それぞれに葉緑体も核も分配される。しかしハテナは片方の娘細胞にしか共生体は受け継がれない。
自然界で採集した試料中に、共生体を持たないハテナが見つかることがある。こういう個体には捕食装置があり、これで藻類を捕らえて共生体とすると思われる。共生体を持つハテナに捕食装置はなく、細胞内の同じ位置は眼点と葉緑体で占められている。共生体を持つハテナと持たないハテナとでは細胞内部の構造が大きく違うのである。
自然界で見つかるハテナのほとんどが共生体を持つ個体であることから、ハテナは生活環の大部分を藻類が共生した状態で過ごすと考えられている。分裂後、共生体を受け継がなかった娘細胞は、捕食装置を発達させて捕食型となり、それがまた1世代のうちに共生体を獲得して捕食装置が退化し光合成型へ変わる、というシステムが確立されていることになる。
共生体を持たない個体が共生体を捕食した後、共生体の核以外の細胞器官は崩壊し、宿主である鞭毛虫の核の支配を受けて葉緑体だけが選択的に増大すると考えられる。共生体はただ葉緑体のサイズを増すだけではなく、宿主の細胞内で特定の位置に配置される。つまり宿主と共生体には、機能的な協調がある程度確立されているのである。しかし宿主の核は共生体の分裂を指示することはできず、分裂の同調はできていない。
このことから、二次共生では宿主と共生体の分裂が同調する前の段階、共生体が葉緑体として確立されるまでの途中の段階があり、ハテナはこの途中段階にあると考えられる。
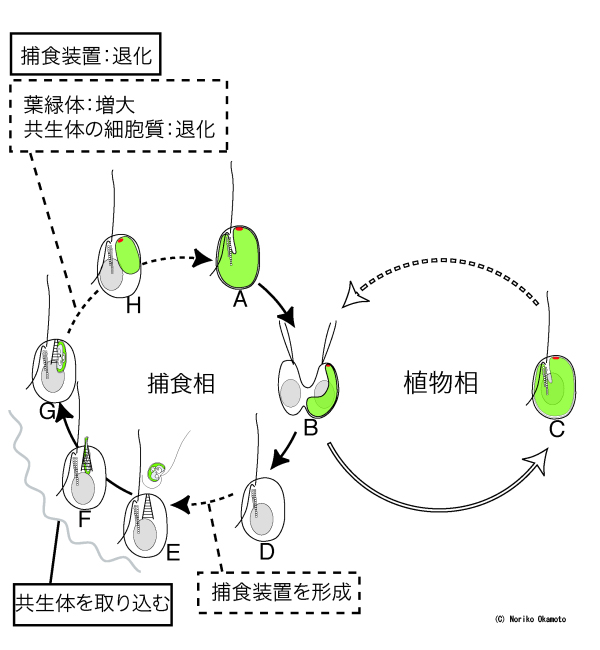
【図5】ハテナの生活環
分裂後、共生体を受け継いだ娘細胞は植物型の生活をし、受け継がなかった無色の娘細胞は捕食型の生活をするようになる。無色の個体は、捕食装置を発達させ、緑色藻類を取り込み共生体とすると考えられる。このような「半藻半獣」ともいえるハテナの生活環は、二次植物の成立過程で、宿主と共生体の分裂が同調する前段階があった可能性を示している。©岡本典子
ハテナは共生体から葉緑体への移行段階を示す。
二次共生とは、元々餌として食べていたものの中から特定のものだけを消化しないで残すようになることだと考えられている。さらに宿主と共生体の細胞周期が同調して初めて、共生体は葉緑体となり、真の植物となり得る。
この従属栄養から独立栄養への過程を、井上教授は「猟師から農夫へ」の転換と例えている。狩猟生活をしていたものが農耕で生きるようになるというのは、非常に大きな決断だ。生きるためのキャリアを変えるためには様々な問題にも直面するだろう。この「植物化」という大変革を遂げる過程で生物が通ったステップを示すものこそ、ハテナだと考えられる。そういう意味で、ハテナの発見は、葉緑体獲得による植物化の過程を明らかにする重要な鍵といえるのである。
井上勲教授へのインタビュー
TJB――ハテナ発見はどのような点が重要でしょうか。
井上――ハテナの生活環は、植物の進化段階の可能性の一つを示しているという点で非常に重要です。現在の植物には多様なグループが含まれますが、その大半は「二次植物」です。二次植物は、光合成をしない無色の生物が真核藻類を取り込む「二次共生」という機構で生まれました。藻類を取り込んだ生物が、真の植物になるには、取り込んだ藻類の細胞周期が宿主の細胞周期と同調し、宿主の分裂時にすべての娘細胞に藻類が受け継がれて、それが継続的に保持される真の「葉緑体」になる必要があります。二次植物の理解のためには、この二次共生の仕組みの解明が非常に重要です。
ハテナはその二次植物になる過程の初期段階、藻類を取り込みある程度の期間存続させてはいるが、分裂周期の同調はまだ確立されていない段階にあるといえます。宿主細胞の分裂時に、共生体は一方の娘細胞だけに受け継がれて、もう一方は再び捕食性生物に戻り、再度、共生体を獲得するといった生活環になっていると思われます。こうした意味で、ハテナの生活環は「半藻半獣モデル」ともいえます。
TJB――ハテナというユニークな名前も注目されています。どういった経緯で命名されたのでしょうか。
井上――正式な学名はHatena arenicola.gen.et.sp.nov. となる予定で、現在申請中です。研究会議での声から、属名を研究室での愛称であった「ハテナ」としました。種小名arenicolaは、ラテン語で「砂の中に住む」という意味で岡本さんが命名しました。なかなか良い学名がついたと思っています。
TJB――藻類研究の面白さや意義をお聞かせください。
井上――30億年かけて今の生命圏を作ってきたのは、藻類です。シアノバクテリア(藍藻)の酸素発生型光合成に始まって、藻類は地球上の生態系を支えてきて、今も支えているという意味で非常に重要な役割を担っています。どのように多種多様な藻類が出現し、多様性を獲得してきたかをきちんと理解することは、藻類に限らず生物界全体の理解にもつながります。
TJB――若い学生へのメッセージをお願いします。
井上――まず、考えるための素材として、本や映画、友達との会話などから「ねた」をたくさん仕入れてほしいと思います。いろいろな考え方を知ることが物事を考える素材となり、素材を蓄積することがとても大切なのです。二つ目は、観察という意味で辺りを見回しながら歩くことです。最後に、冒険をすることです。考えすぎてもチャンスを逃してしまいます。これだと感じたら、躊躇せずに飛び込むことが大事です。新しい自分の世界を、自分で切り開いていってほしいと思います。

藻類の多様性について、電子顕微鏡写真を用いて解説される井上教授
筑波大学大学院生命環境科学研究科長。現在の研究テーマは、藻類の多様性。 生物学類では2・3年次の植物系統分類学Ⅰなどの講義を担当。 藻類の分類を通じて植物界全体の進化の解明を目指している。 30億年という時の流れの中で、生命、地球がどう進化してきたかを藻類というユニークな視点から捉えた著書『藻類30億年の自然史』 (東海大学出版社刊)が2006年1月に出版された。
ハテナ発見者、岡本典子氏へのインタビュー
TJB――ハテナ発見の経緯を聞かせてください。
岡本――2000年11月末に、和歌山での学会に参加した時に砂浜で採集した試料(砂)から偶然発見しました。井上研では新しい生き物を探すための採集が皆の習性で、私も出かけた先で必ずやりました。この時も同様で、研究室に戻ってからやっとその試料を顕微鏡で見るとなんと鞭毛で容器の底をぺたぺたと這い、微細藻類にしては大きめの0.03mmほどの生き物がいたのです。どの藻類とも違うので「ハテナ虫」と呼び始めました。
TJB――分裂細胞はどのように発見されましたか。
岡本――ハテナは研究室では培養できないので、細胞は現地調達していました。採集してきた細胞は弱っていくだけなので、必死で顕微鏡に向かっていたところ、夜半から朝方に分裂細胞を発見したのです。この時間帯は藻類の細胞分裂が活発です。
TJB――井上研究室を選んだ理由をお聞かせください。
岡本――井上研究室の新しい生き物を見つけて研究するという伝統に魅力を感じたからです。また、大学2年時に臨海実習で新しい珪藻を発見していたというきっかけもありました。生き物の秘密を「見る」ためには人間が生き物に寄り添い、見せてくれるものを受け止めようとします(そのため最後には、なんと研究者と生き物は似てくるのです!)。この種の研究はすべて本人の「自由」で進められます。大学の教育を(高校の延長のように)受け身にとらえている人にとっては「自由」研究を行うというのは、目標が曖昧で難しいことだと思いますが、私は生き物を「見る」ことも、未知の領域に入っていくことも好きだったため、井上研での研究が合っていたのです。
TJB――「サイエンス」に論文が掲載された感想はいかがですか。
岡本――科学雑誌の中でも「サイエンス」と「ネイチャー」は、一般の人でも楽しめる内容が取り上げられる娯楽的要素のある雑誌です。読者数が厚いので、掲載されると知名度も上がります。日本ではニュースでも取り上げられ、多くの人に知ってもらえたようで、うれしいです。
TJB――今後の研究目標をお聞かせください。
岡本――ハテナの研究で、共生体が葉緑体になる過程に興味を惹かれました。その現象を調べる上で必要な分子生物学的手法を学ぶため、メルボルン大学へ来たのです。日本の大学とは違うところが多く、私はいろいろな価値観に触れています。
ここではマラリア原虫の研究をしています。これはヒトの血液に寄生する、実は二次共生由来の退化した葉緑体を持つ「元・植物」です。全ゲノム配列解析や遺伝子導入の系が整備された研究基盤があるため、これを「モデル生物」とし、細胞分裂時の葉緑体と細胞骨格間の相互関係を調べています。将来的には、このような実験系をハテナのような葉緑体獲得段階にある生物で確立するのが目標です。

岡本典子氏
2000年筑波大学生物学類卒業。2005年同大学生命環境科学研究科卒業。理学博士。現在は、日本学術振興会特別研究員としてオーストラリアのメルボルン大学McFadden研究室に所属。共生体が葉緑体になる過程の解明を目指して、マラリア原虫を材料として研究を行っている。写真は、国際藻類学会で訪れた南アフリカ共和国にて。
Communicated by Shinobu Satoh, Received January 30, 2006. Revised version received March 3, 2006.
©2006 筑波大学生物学類
|