|
つくば生物ジャーナル Tsukuba Journal of Biology (2006) 5: TJB200609TM.
動物系統分類学 ―私の昔のテキストから―(連載第2回)
牧岡 俊樹(元 筑波大学 生物科学系)
第1章 生物の分類と系統(続き)
第2項 進化と系統
系統 (phyletic line) とは進化の足跡をつなぎ合わせた道筋である。現在に至る地球上の生物の進化の歴史は1度限りのことであるから、真実の系統はただ1つしかないが、われわれは時間を逆行して直接それを見ることも、実験的に再現して見ることもできない。ゆえに現在いろいろ提案されている生物界の系統関係
(phylogenetic relationship) はすべて、生物の表現型(形や機能)や遺伝子型(DNA またはその転写産物である RNA
の塩基配列)に関する乏しいデータを寄せ集めて推定されたものであり、真実の系統を限られた場所や方向から見たスケッチのようなものである。
2─1 自然分類体系と系統分類学
理想的な分類体系 (classification system) は、1度限りの進化の歴史にもとづく唯一の自然分類体系(natural c.
s.)である。すべての生物種は類縁 (affinity) の近い順(種分化の順)に位置し、類縁が遠くなれば次第に上位の上種分類群に至る。また種分化(進化)の歴史を順にさかのぼって行くと次第に古い祖先(ancestor)に到達し、最終的には生命の起源に至る。進化の歴史の結果としてこのような自然分類体系の存在することは明らかであるが、現在のわれわれは進化の歴史を直接見ることはできず、したがってその結果である生物種間の類縁、つまり自然分類体系の実像を直接知ることができない。それを近似的に知るための方法が系統分類学
(systematics, phylogenetic s.) であり、種を基礎としていろいろな分類群の範囲を明らかにする分類学(taxonomy)と、分類群間の進化的類縁を明らかにする系統学(phylogeny)を合わせたものである。系統分類学の基本的な方法は、眼に見ることのできる生物の表現型や遺伝子型の特徴を比較して、眼に見ることのできない類縁の遠近を推定することである。
だが現状では、系統分類学の基礎となるべき種のほとんどが形態種の段階にあり、また種間の類縁を推定するための生物の表現型のデータは客観的に比較できるものが少なく(本章2-2)、遺伝子型のデータはまだ数も種類も乏しく十分な比較ができない状態にあるので、現在の分類体系そのものに多くの不明の点や不自然な部分が残り、理想的な自然分類体系からは遠い段階にあると言わねばならない。不明の点を明らかにし、不自然な部分を修正して、真の自然分類体系に少しずつでも近づけることが現在の系統分類学の課題であり、そのためには分類学の分野だけでなく、生物学の全分野からの新しい情報が待たれている。そしてそれによる分類体系の進歩(自然分類体系への接近)は、生物学の全分野にとって必要なことである。
|
コラム7:ただ1度とただ1つ
地球上での生命の起源は必ずしも1度ではなかったかもしれないが、現在の生物につながる地球上での生物の進化はただ1度であったと信じられる。その最も本質的な根拠は全生物界に共通に見られる DNA の4種類の塩基3個ずつの64通りの組み合わせとタンパク質の20種類のアミノ酸の配列との対応関係、いわゆる普遍暗号(universal code)の存在である。原核生物(第2章1-1)からわれわれ真核生物(第2章1-2)に至るまで、調べられた限りの生物で基本的に同じ対応関係が用いられている(少数の生物にごく部分的な対応関係の違いがあるが、それらは後から変化したものと思われる)という事実は、約38億年の生物進化の歴史を通じて維持され続けたこの対応関係の信じがたいほどの高い安定性を示していると同時に、このような密接で複雑な対応関係が独立に複数回発生したとは確率的にも考えられないことから、この対応関係はただ1つの祖先生物においてただ1度生じたのであり、ゆえに現在に至る全生物界の進化の歴史はただ1つであることを示している。
|
2─2 相同と相似
生物の種は時間とともに変異を蓄積し、種分化を重ねてしだいに類縁の遠い種に、さらに上種分類群に至ったと考えられるので、現時点での分類群間の形や性質など表現型上の特徴(形質:character)あるいは遺伝子型(DNA またはその転写産物である RNA の塩基配列)上の特徴を比較すると、よく似ているほど類縁が近く、似ていないほど類縁が遠いと考えられる。しかし、似ている特徴にも、親子兄弟姉妹のように類縁が近いための類似と、他人のそら似のように類縁に関係のない見かけ上の類似とがある。また、類縁の近い親子兄弟の間でも、形質ごとに類似の程度は同じではない。
2-2-1 相同
類縁の近さを反映した類似とは、たとえば人類と猿類の平面的で毛の少ない顔や、平爪の生えた5本の指のうち母指と他の4本が向き合っている(対向母指)手のように、共通祖先に由来する共通起源の形質、つまり相同(homology)と呼ばれる形質の類似である。だが、同じ相同の形質にも、たとえば人類と猿類の足の指のように、平爪の生えた5本の指が、猿類では手と同じく対向母指であるが人類では5本の指がみな同列に並んでいるように、あまり似ていないものもある。これは、猿類は樹上で生活し、足の指も手の指と同じく枝をつかむので、ともに対向母指である方がよいが、人類は地上で直立二足歩行をするので、足の指は5本並んで地面を強く踏むようになったものと思われる。このように、表現型の相同の形質には類縁を反映した類似の他に、生活様式や機能の類似や差異の影響が含まれ、その影響の種類や程度は形質によって異なるので、相同の形質の類似によって類縁を判定する場合には、生活様式や機能の影響のなるべく少ない形質を選ぶ必要がある。また類似の差異を平均化するために、なるべく多くのそのような形質を比較してみる必要がある。
遺伝子型の特徴でも、ある遺伝子またはその転写産物の塩基配列を比較して分類群間の類縁を判定するには、必ず共通の祖先遺伝子から由来した相同の遺伝子またはその転写産物の塩基配列を比較しなければならない。たとえば、すべての真核生物の細胞にあるタンパク質合成装置である80Sのリボソーム(Sは分子の大きさの単位。原核生物のリボソームは70S)を構成するいろいろな RNA のうち、18SリボソームRNA(18S-rRNA)またはその相補的 DNA(18SrDNA)は分類群間の類縁の判定に、相同の塩基配列としてよく用いられる。しかし遺伝子型に生じる突然変異は、DNA 全体としてはほぼ一定の頻度で生じるとしても、生じる位置はランダムであるから、どの遺伝子にも同じ頻度で生じているとは限らない。ゆえになるべく多くの相同の遺伝子の塩基配列を比較して類縁の判定をすることが望ましいが、多くの分類群間で比較できる相同の遺伝子またはそれらの転写産物の種類はまだ少ない。
2-2-2 相似
類縁の遠い(近縁の共通祖先がない)分類群の間でも、生活様式や機能が似ているために形態など表現型の形質がよく似ている場合がある(たとえば軟体動物門頭足綱のカメラ眼と脊索動物門脊椎亜門のカメラ眼の類似など)。このような共通起源でない(相同でない)形質が似ていることを収斂(convergence)と呼び、収斂の結果である類似を相似(analogy)と呼ぶが、相似による類似は系統上の類縁を反映していないので、系統関係推定の根拠とすることはできない。また遺伝子型では、進化的出発点としての共通の祖先遺伝子から由来していない(相同でない)遺伝子の DNA またはその転写産物である RNA の塩基配列の類似の程度は、系統関係を反映していないので、系統関係推定の根拠とすることはできない。
|
コラム8:表現型と遺伝子型による系統解析
表現型では、相同の形質は形や性質など種類が多く観察もしやすいが、その多くは環境からの自然選択の作用を直接受けるので、系統の遠近以外に環境への適応度の大小など別の要素が混入しやすく、相同の形質の種類によって類似の程度は必ずしも同じではない(本章2-2)。また類似の程度を客観的に判定する(数値化する)こともむずかしく、表現型の比較には研究者の主観が入りやすいとよく言われる。だが分子生物学の発展以前には、系統解析に使える生物の特徴は表現型しかなかったので、多くの研究者がそれまでの研究で蓄積された多数の相同の形質の中から系統関係以外の要素の混入が少ないと思われる形質(体腔や排出器官など体内の形質や胚発生中の形質など)を選んで系統関係を考察してきた。本講の採用する系統関係もその1つであるが、近年の遺伝子型にもとづく系統関係との間にはいろいろな不一致点が存在する(第2章2-4)。
遺伝子型は、すべての表現型の元となる一次データであり、それにもとづいて推定される系統関係はより信頼性が高いと思われる。しかし遺伝子型は研究の歴史が浅いので、生物群間で広く相同性が認められている遺伝子または遺伝子産物のデータはまだ少なく、特に綱や門のような大きな分類群の間では、ごく限られた遺伝子または遺伝子産物の比較しかできない現状である。また遺伝子や遺伝子産物のデータの得られている生物種もまだわずかで、このため1種か2種のデータで1つの綱や門を代表することも多いが、これでは綱や門の中の多様性を十分に反映しているとは言えない。また、表現型から想定された綱や門のような分類群が、遺伝子型からも同様に成立するのかどうかの検証もできない。
遺伝子型の今後の課題は、多種類の相同の遺伝子または遺伝子産物のデータを得ることと、それらのデータの得られている生物種の数を増すことにあると思われ、それによって、推定される系統関係の信頼性の向上とともに、表現型にもとづく系統関係とのさまざまな不一致の原因の解明が期待される。
|
2─3 系統図(系統樹)
ラマルク(Lamarck,1809)は動物群間の進化にもとづく系統関係を、動物群間を線でつないで初めて図示した(図1)。またヘッケル(Haeckel, 1866) は、共通の祖先から複数の子孫が次々に分岐して生じた進化の過程を枝分かれした樹の形で表現した(図2)。ヘッケルの図は多くの分類群間の複雑な系統関係を一見して理解できるわかりやすさがあるが、まだわかっていない部分までわかっているかのように枝でつないでしまう欠点がある。その後、系統分類学の発展とともに多くの系統図が提案されたが、ヘッケル風の系統樹(phylogenetic tree) よりもラマルク風の線状の系統図 (phylogenetic diagram) が広く用いられたのは、描画が簡単なこととともに、樹の形にとらわれず、不確かな部分は点線でつなぐなど表現の幅が広いためと思われる。なお、線状の系統図でも、枝分かれしているので系統樹と呼ぶことがある。
ヘニッヒ(Hennig, 1950)以来の分岐論(cladistics)は、ヘッケルの系統樹の枝の分岐と分類群の関係をより理論的に定義した。現生の種はみな、それぞれの祖先種から種分化(分岐)をくりかえして進化して来たので(図3-1)、共通の祖先種から生じた全部の種は、その祖先種が初めてもった形質を最新の相同の形質(共有子孫形質、共有派生形質 synapomorphy)として受継いでいる。ゆえにそれらの種は、単一の共通祖先の子孫である単系統群(monophyletic group)であり、属などの上種分類群として認められる。しかし、単一の共通祖先から生じた単系統群の一部の種(側系統群 paraphyletic group)や、近くに共通祖先を想定できない種同士(多系統群 polyphyletic group)は、独自の分類群とは認められない。
分岐論にもとづく分岐図(cladogram)は、系統図を客観的に描くための方法の1つとして広く使用されるようになった。この理論的な方法によって、たとえば現生の陸生脊椎動物の分岐図(図3-2)のように、それぞれの単系統群が明確に示されるようになったが、その結果、従来の爬虫類が側系統群となり、分類群として成立しなくなるなど、今までの系統分類体系と一致しない点も生じた。しかし分岐論による分類は、客観的で数値化しやすく、比較しやすい長所があり、いろいろなコンピュータプログラム(最節約法、最尤法、近隣結合法など)が考案され、データにもとづいていくつもの系統図が比較的容易に描けるようになった。
現在の地球上のすべての生物が唯一の共通祖先から出たものだとすれば、真の系統図はただ1つしかあり得ないが、それを正しく描くには、各生物群についての現在の知識はまだあまりに乏しく、したがって現在のコンピュータプログラムも、少ないデータから矛盾の少ない系統関係を導き出すために、さまざまな仮定を含んでいる。これからは、信頼性の高い表現型や遺伝子型のデータを多く得ることによって、多数の分類群間で系統関係を反映する多数の相同の特徴を比較できるようにすることが最も重要であり、それらの多数のデータの比較を厳密に客観的に行うすぐれたプログラムの開発が求められる。
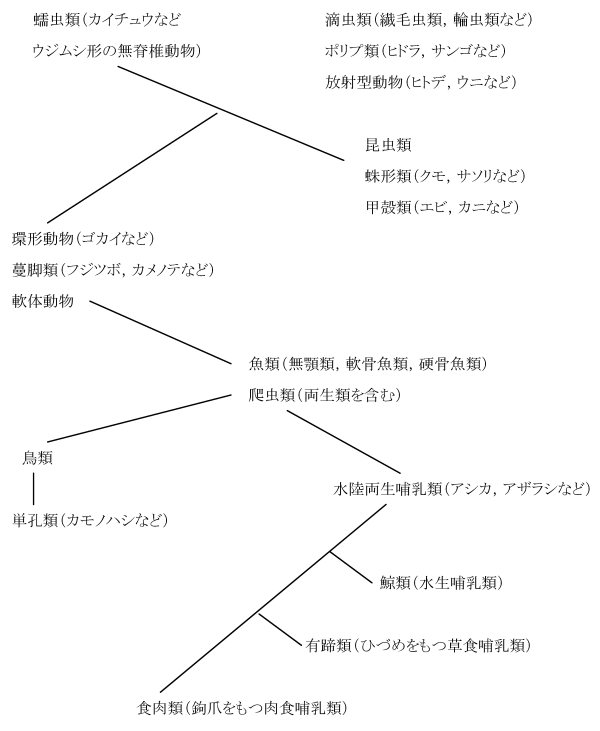
図1:ラマルク(1809)による最初の動物系統図
蠕虫(ぜんちゅう)類から昆虫類などの節足動物が生じ、その途中から環形動物と軟体動物が分かれた。甲殻類である蔓脚(まんきゃく)類は当時は軟体動物に近いと思われていた。軟体動物から魚類と爬虫類(当時は両生類を含んでいた)が生じ、以後は脊椎動物になる。爬虫類から左側には鳥類が生じ、鳥類からさらにカモノハシが生じているのは、当時ヨーロッパに標本がもたらされて間もなかったカモノハシが哺乳類であることはまだわかっていなかったためである。また右側にはまず水陸両生の哺乳類が生じ、続いて水生の鯨類、さらに陸生の草食哺乳類と肉食哺乳類が生じている。なお右上の滴虫類、ポリプ類、放射型動物は系統関係が明確でないとしてあえて線でつながなかった。
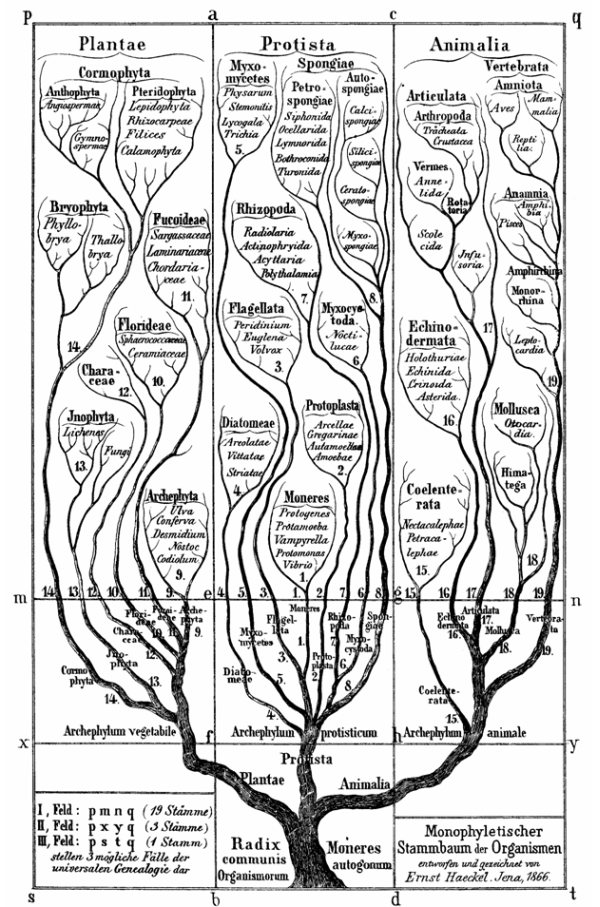
図2:ヘッケル(1866)の系統樹
生物界全体の単系統性(Monophyleticity)を示している。
全生物の共通祖先である1本の幹から植物(Plantae)、原生生物(Protista)、動物(Animalia)の3つの枝が分かれ、それぞれが次々に分岐して多数の分類群を生じる。分類群間の類縁は分岐点(分類群間の共通祖先)からの分岐の回数が多いほど遠い。
|
コラム9:系統図の方向と進化の方向
ラマルク(1809)の図では進化の方向は上から下に向かっているが、これは進化論以前のキリスト教的世界観にもとづく自然の体系(神の創造物のうち、下等なものを下に、高等なものを上に置き、人間を動物界の最上位とする体系)を逆転させた(ただしこの図には人間は含まれていないが)きわめて革新的なものである。だが、ダーウイン(Darwin, 1859) の種の起源の中の有名な進化の概念図(系統図ではない)が下から上に向かって描かれてから後、系統図はみな下から上に向かうようになり、これが古い自然の体系の方向と一致したため、高等・下等の概念がよみがえり、向上的進化の視点が導入されることとなった。
だが現在のどんな簡単な生物でも、むしろ簡単な生物であるほど、われわれ複雑な多細胞生物よりもはるかに長い進化の歴史を生きてきたのであり、ゆえにわれわれよりも「進化を重ねた」生物だと言える。生物の進化には、多細胞体の複雑さ以外にも、単細胞としての洗練や、少細胞での可塑性や、寄生にともなう体制の単純化と生活史の複雑化などじつに多くの方向があり、それぞれの方向の成功者が現在の生物界の多様性を作っているのである。
なお、近年の分岐論にもとづきコンピュータによって描かれる系統図には左から右に向かう横向きのものが多い。ここでは分岐の有無と順序が重要で、分岐の確からしさを%で表示することもできる。分岐の結果生じた各分類群を示す横線はふつう平行に描かれ、進化の方向性は考慮されないが、横線の長さで分岐の時点以後の相対的な時間の長さを示すこともできる。
|
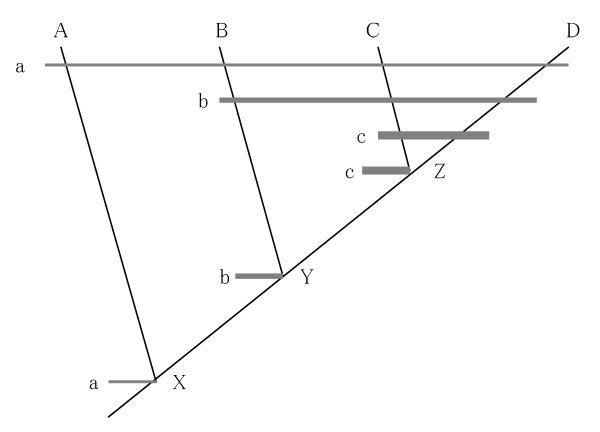
図3-1:種 A, B, C, D の分岐図
現生の種 A、B、C、D はそれぞれ仮想の祖先種 X、Y、Z の分岐によって順に生じ、祖先種 Y は X から、Z は Y からそれぞれ分岐して生じたものとする。また形質 a、b、c は祖先種 X、Y、Z でそれぞれ生じ、相同の形質としてそれぞれの子孫種に伝えられているものとする。
現生の種群 A-D は相同の形質 a を、B-D は相同の形質 b を、C-D は相同の形質 c をそれぞれの祖先種X、Y、Z から来た子孫形質(派生形質 apomorphy)として共有するので、種群 A-D、B-D、C-D はそれぞれの祖先種 X、Y、Z から出た最も近縁な種群(単系統群 monophyletic group)であり、独自の上種分類群 A-D、B-D、C-D としてまとめることができる。なおこの場合、C と D は Z を共通祖先とする姉妹種、B と C-D は Y を共通祖先とする姉妹群、A と B-D は X を共通祖先とする姉妹群であり、C-D は B-D の、B-D は A-D のそれぞれ下位の分類群である。
一方、種群 A-B と A-C は形質 a を共有し、種群 B-C は形質 a、b を共有しているが、形質 a、b は他に種 D も共有している。したがって、種 D を含まない種群 A-B、A-C、B-C はいずれも単系統群ではなく、近縁な種群の一部(側系統群 paraphyletic group)にすぎないので、独自の上種分類群としてまとめることはできない。なお、このような分岐図はたとえば次の図(図3-2)のように横向きにも描くことができる。
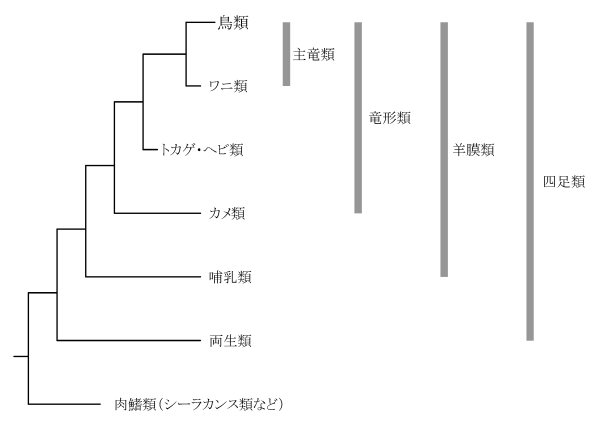
図3-2:現生の陸生脊椎動物グループの模式的な分岐図
両生類から鳥類までの各グループはそれぞれの祖先から由来した単系統群であり、また全体としては肉鰭(にくき)類と姉妹群となる単系統群(四足類)である。またワニ類と鳥類はトカゲ・ヘビ類を姉妹群とする単系統群(主竜類)としてまとめられる。カメ類から鳥類までのグループは哺乳類と姉妹群になる単系統群(竜形類)として、哺乳類から鳥類までのグループは両生類と姉妹群になる単系統群(羊膜類)としてそれぞれまとめられる。しかしカメ類とトカゲ・ヘビ類とワニ類のグループは単系統群である竜形類の一部(鳥類を含まない)であり、したがって側系統群であるから、分岐論では独自の分類群(爬虫類)としてまとめることができない。(本図は模式的な図なので分岐の確からしさの指標(%)の数値は表示していない。また横線の長さは分岐の時点からの相対的な時間の長さを特に考慮していない。)
ーー次回に続くーー
Contributed by Toshiki Makioka, Received September 4, 2006.
©2006 筑波大学生物学類
|