|
つくば生物ジャーナル Tsukuba Journal of Biology (2007) 6: TJB200701200310740
テトラヒメナにおけるアクチン細胞骨格制御因子、Arp2/3タンパク質複合体の研究
生方 寛昭 (筑波大学 生物学類 4年) 指導教員:沼田 治 (筑波大学 生命環境科学研究科)
【目的・背景】
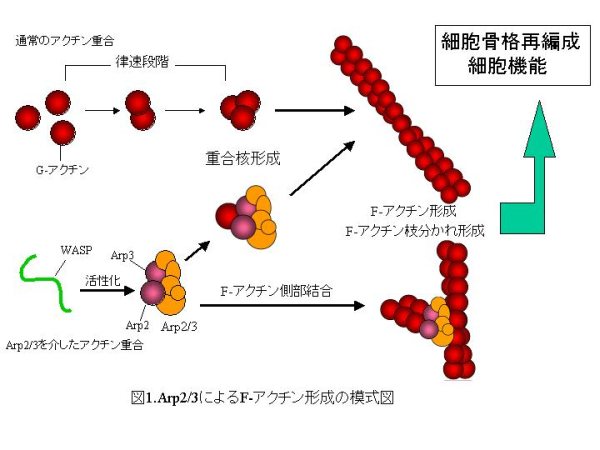
アクチン細胞骨格は単量体のG-アクチンが重合し、F-アクチンとなることで形成される。アクチンが重合を開始するには三つのG-アクチンが重合した重合核の形成が必要であり、これがF-アクチン形成の律速段階となっている。二つのアクチン関連タンパク質(actin related protein ; Arp)Arp2及びArp3を含むヘテロ七量体のタンパク質複合体Arp2/3はアクチン重合核を模倣して、律速段階をスキップしたアクチン重合を誘導する。この機能は細胞内での細胞骨格の素早い再編成に重要である。さらに、Arp2/3が活性を発揮するには、既存のF-アクチンの側部に結合し、WASPなどの活性化因子からシグナルを受けることが必要である。この結果、Arp2/3を介して重合したF-アクチンは枝分かれする。この仕組みは、内外のシグナルに対応して細胞骨格の局所的な再編成を行うために非常に重要である(図1)。
Arp2/3が細胞機能に果たす役割は様々な生物で調べられており、これまでに細胞の遊走、ファゴサイトーシスやエンドサイトーシス、小胞輸送などに関わることが知られてきた。細胞質分裂について見ると、ショウジョウバエや線虫を用いた実験ではArp2/3は不要なことが示されているが、分裂酵母を用いた実験ではArp2/3の関与が示唆されている。
繊毛虫テトラヒメナは細胞の前方にある口部装置で頻繁にファゴサイトーシスを行う。これにより取り込まれた物質は、細胞内へ漏斗状に伸びたディープファイバーを伝わって食胞を形成する。また、テトラヒメナは動物細胞と同様に収縮環を利用した細胞質分裂を行なう。動物細胞や分裂酵母などのArp2/3の機能が調べられてきた細胞と、テトラヒメナは大きな進化的距離がある。そのため、テトラヒメナのファゴサイト―シスや細胞質分裂におけるArp2/3の役割は、生物の進化を考える上で非常に興味深い問題である。また、繊毛虫と近い生物群であるアピコンプレクサ類(マラリアやトキソプラズマなどの寄生原虫)は明確なArp2/3を保持していない。しかし、それに対応するアクチン調節因子を持つ可能性がある。そのため、テトラヒメナの細胞機能についての知見は、寄生原虫の性状を知る上で有用だと考えられる。以上の理由から、私はテトラヒメナのArp2/3について研究した。
【方法】
1.cDNAのクローニング
Arp2/3の構成サブユニット遺伝子と、Arp2/3の活性化因子WASP遺伝子の全長配列をクローニングし、塩基配列を決定した。
2.Arp2/3構成サブユニットの系統解析
動物、植物、粘菌、アピコンプレクサを含む9種の生物種で得られるArp2/3構成サブユニットのアミノ酸配列情報を集め、これにテトラヒメナの配列を加え、最適アラインメントを基に無根系統樹を作成した。
3. WASP及びArp2/3サブユニットARPC3に対する抗体の作成
抗WASP抗体及び抗ARPC3抗体を得るために、WASPのタンパク質断片(267〜419a.a.)及びARPC3の全長配列(1〜158a.a.)をGST融合タンパク質として大腸菌に発現させた。発現したタンパク質を精製し、アジュバンドと混合し、エマルジョンとしたものをウサギに皮下注射した。
得られた抗血清はHis-タグをつけて発現したWASP断片及びARPC3全長配列を用いて、アフィニティー精製した。
4.間接蛍光抗体法によるテトラヒメナの免疫染色
テトラヒメナのARPC3の局在を観察するために、免疫染色を行った。細胞を固定後、抗体を反応させた。その後、蛍光標識二次抗体を反応させ、蛍光顕微鏡で観察した。
【結果・考察】
Arp2/3サブユニットの無根系統樹から推定される分子の類縁関係は、進化系統樹から考えられる生物の分類によく一致することが分かった。このことは、Arp2/3が生物進化の過程で機能を保ちながら、緩やかな変化を受けてきたことを意味する。一方、ARPC5は、他の構成成分と較べるとその保存性が低いことが分かった。ヒトや出芽酵母のARPC5はArp2/3の重合核機能に強い影響を持っているにも関わらず、進化の過程でアミノ酸がよく置換していることは興味深い。もしかしたら、それぞれの生物(細胞)の性質に合わせてARPC5が変化することがアクチン細胞骨格の制御に大切であったのかもしれない。
アフィニティー精製したARPC3抗体を用いてテトラヒメナの抽出液に対するウエスタンブロッティングを行ったところ、ARPC3の予想分子量と一致するバンドが認識された。そこで、ARPC3抗体を使って細胞を免疫染色した結果、口部装置とディープファイバーに局在が見られた。このことから、Arp2/3はテトラヒメナにおいて食胞の形成に関わることが推測された。
【今後の予定】
今後は、抗Arp2抗体も作成し、より詳細にArp2/3とWASPの局在を調べる。さらに、各遺伝子をノックダウンし、細胞における機能解析を行なう。また、WASPによるArp2/3のアクチン重合能の活性化について検討を行なう。
©2007 筑波大学生物学類
|