|
つくば生物ジャーナル Tsukuba Journal of Biology (2007) 6: TJB200701200310790
テトラヒメナの膜輸送に関わるRabタンパク質の研究 ― 私の軌跡(奇蹟)
中原 徹 (筑波大学 生物学類 4年) 指導教員:沼田 治 (筑波大学 生命環境科学研究科)
【目的・背景】
地球上に存在する生物は、動物、植物、菌類、プロティスタからなる真核生物と、古細菌と真正細菌からなる原核生物に大別される。前者と後者の最大の差は、核膜をはじめとする細胞内膜系の有無にあるといっても過言ではない。真核生物の細胞内膜系は、核、小胞体、ゴルジ体、リソソームなどのオルガネラの機能を特徴付け、互いに独自性を発揮している。しかし、その一方で各オルガネラ間は膜輸送などを介して相互作用することが必要である。
いかにして細胞内膜系が相互作用しつつ、独自性を発揮できるのであろうか?この問いに対する解答の1つが、Rabファミリー低分子量Gタンパク質(以下Rab)の働きである。1980年Schekmanらによって細胞内小胞輸送の必須遺伝子の1つSEC4が出芽酵母の遺伝学的解析から単離され、後にこれがRabファミリーの1つであり、ゴルジ体−細胞膜間の小胞輸送の制御を担うことが明らかにされた。1992年Zerialらが初期エンドサイトーシス経路でのRab5の働きを明らかにした。2000年、彼らはアフィニティークロマトグラフィーによって、GTP結合型Rab5に22種類ものタンパク質が特異的に結合することを明らかにした。現在、細胞内には複数のRabが発現し、それぞれが細胞内の特定の膜へ局在し、その部位で膜の会合及び輸送制御を行っていると考えられる。
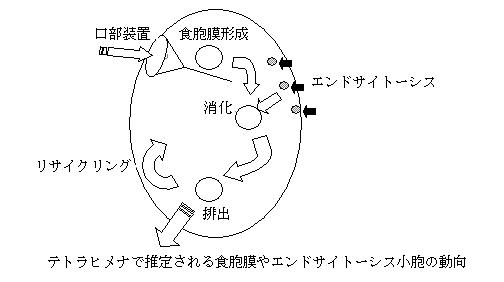
繊毛虫Tetrahymena thermophilaは、上述したオルガネラに加えて、アルベオラーサックや食胞などの細胞内膜系が発達している。細胞前方の口部装置から活発に物質を食胞に取込み、食胞は内容物を消化しながら輸送され、後方の細胞肛門から未消化物を排出し、食胞膜はリサイクリングされる。また、テトラヒメナは小胞輸送の仕組みがよく調べられている動物や菌類と異なるプロティスタに属し、その小胞輸送の仕組みを知ることが、生物進化における真核生物の起源を知るヒントになると思われる。興味深いことに、T. thermophilaにはRab遺伝子が、64種存在することが報告されており、この数はヒトに匹敵している。
今回、私は、食胞形成やエンドサイトーシスなどの外来物質を取り込む一連の流れを解析するために、Rab5の解析を行った。さらに、細胞内に取り込まれた膜のリサイクリングを制御すると推定されるRab11の研究にも着手した。
【実験】
1.cDNAのクローニング
T.thermophila のcDNAライブラリーより、Rab5様遺伝子群のRab5A、Rab5B、Rab5C、Rab17A、Rab21A、Rab22の全6種、及びRab11A、Rab11B、Rab11C、Rab11D、Rab11E、Rab11F、Rab11Gの7種をPCR法により増幅し、クローニングした。DNAシークエンスを行い、塩基配列を調べた。
2.大腸菌を用いたRab5A全長配列の発現と、抗体の作製
Rab5Aの cDNA全長配列をpGEX5X-1に挿入し、大腸菌に導入した。T. thermophilaのグルタミンをコードするTAAやTAGは、大腸菌を含めた一般的生物では終止コドンとなるため、CAA及びCAGに改変した。発現誘導を行った大腸菌を破砕し、回収したGST融合タンパクとアジュバンドを混合したエマルジョンを、ウサギに皮下注射した。
3.血清のアフィニティー精製
Rab5Aの cDNAをpET28c(+)に挿入し、大腸菌内で、His-Rab5Aを発現させた。免疫したウサギの抗血清を、精製したHis-Rab5Aを用いてアフィニティー精製し、抗Rab5A抗体を得た。
4.間接蛍光抗体法によるテトラヒメナの観察
T. thermophila をメタノール固定後、Rab5A抗体と反応させた。洗浄後、蛍光標識抗ウサギIgG抗体と反応させた。余分な抗体を洗浄により除去し、蛍光顕微鏡で局在を観察した。
【結果・考察】
cDNAをクローニングした結果、Rab5様遺伝子群の6種類全てと、7種類のRab11が発現していることを確認した。これらの内、一部のものはゲノムプロジェクトで予想されるエキソンとイントロン配列と一致しないことが判明した。そのため、新たにRab遺伝子群の系統解析を試みた。
免疫したウサギの血清によるウェスタンブロッティングの結果、Rab5Aの予想分子量である22kDaにバンドが検出された。アフィニティー精製した抗Rab5A抗体を用いて、免疫染色を試みた結果、細胞全体にドット状の局在と、口部装置への局在が見られた。細胞全体でのドット状の局在は、エンドサイトーシス小胞と考えられた。一方、口部装置への局在は、細胞肛門で未消化物を排出した食胞膜が、口部装置にリサイクリングされた状態を反映していると考えられた。
今後、抗Rab5A抗体を用いた分画実験やアンチセンスrRNA法を用いたRab5Aのノックダウンによる機能解析を行いたい。さらに、Rab11の機能解析も進めていきたいと思う。
©2007 筑波大学生物学類
|