|
つくば生物ジャーナル Tsukuba Journal of Biology (2007) 6: TJB200701200310805
円石藻Emiliania huxleyiにおける長鎖不飽和脂質アルケノン合成酵素遺伝子の探索
安賀 愛 (筑波大学 生物学類 4年) 指導教員:白岩 善博(筑波大学 生命環境科学研究科)
アルケノンは、ハプト植物門の中でもEmiliania huxleyiやGephyrocapsa oceanica, Isochrysis galbana,およびChrysotila lamellosa のみによって合成される特徴的な不飽和ケトンで、C37からC39の非常に長い炭素鎖を持つ。また、生体内の一般的な脂質は不飽和結合がcis型で酸化・分解を受けやすいのに対し、アルケノンの不飽和結合は全てtrans型であるため、非常に安定で堆積物として残りやすい。更に、この脂質の不飽和度から計算されるアルケノン不飽和指数 (Uk’37) は生合成時の温度を正確に反映することから、地球科学の分野で地層形成時の海洋温度を推定する強力な古水温計として用いられている。(Prahl et al., 1988) アルケノン不飽和指数(Uk'37)
しかしながら、アルケノンの代謝経路や生理的機能などの生理学的情報は少なく、不明な点が多い。したがって、アルケノンの合成および代謝系の解明は新規脂質代謝経路の発見に繋がり、アルケノン古水温計の精度を高めるのにも重要な知見を与えるものである。よって、本研究はアルケノン合成系の解明を目的とした。
先行研究から、脂肪酸のde novo合成のKey酵素であるβ-ケトアシルACPシンターゼ(以下、KAS)反応を阻害するセルレニンを培地に添加すると、アルケノンの合成が著しく低下することが分かっており(Shiraiwa et al. 2005)、アルケノンは脂肪酸合成系を経ている可能性が高い。一般に、生体内の長鎖脂肪酸はde novo合成系でC18まで伸長した後、更に別の伸長反応経路を経て合成される。この伸長反応は、動物や酵母型のElongase(ELO)と、植物型のFatty Acid Elongase(FAE)という膜結合型脂肪酸縮合酵素によって触媒される。植物はこの両方を有するが、動物や酵母はELOしか持たない。(Trenkamp et al.,2004)
KAS、ELOおよびFAE全ての酵素に相同性の高い配列がEmiliania ESTデータベースで見つかったことから、それらの酵素がE. huxleyiにおけるアルケノン合成に関与しているか否かを確認することを試みた。そのために、アルケノンの合成が低温条件で促進される (Shiraiwa et al. 2005)ことを利用し、まず、低温誘導下でそれら遺伝子の発現解析を行った。
【結果・考察】
E. huxleyiのESTデータベースではシロイヌノナズナのKASと相同性のある配列が3種、出芽酵母のELOと相同性がある配列が3種、シロイヌノナズナのFAEと相同性のある配列が1種見つかった。そのうちKASと相同性のある配列一種を除く全てをクローニングした。以下、それら配列を、EhKasα、EhKasβ、EhEloα、EhEloβ、EhEloγ、EhFaeとした。
E. huxleyi細胞は前培養として2日間、20℃で静置培養し、OD750が0.1になった時に、10℃の水槽に移した(20℃のまま培養を続けたものをコントロールとした)。低温誘導開始後3日目までの細胞試料からRNAを回収し、ノーザンブロッティングによる発現解析を行った。培養時の光強度は100μmolm-2s-1に調節した。
その結果、EhKasα、EhEloα、およびEhEloγの発現が低温ストレス下で顕著に増加した。EhEloβおよびEhFaeでは発現レベルの変化は見られず、EhKasβではシグナルの確認ができなかった。
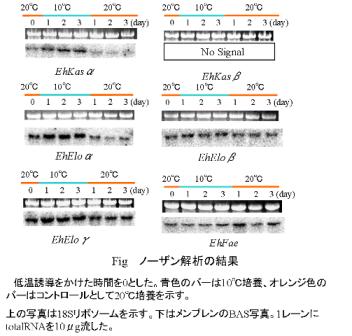 以上の結果からEhKasα、EhEloα、EhEloγは低温誘導性であることにおいて、アルケノン合成との何らかの関与が考えられる。一方、EhEloβとEhFaeは低温誘導を受けず、アルケノンの合成には直接関与する可能性が低いと考えられる。
【今後の予定】 1.アルケノン合成を誘導する要因の探索 本結果だけで目的の遺伝子がアルケノン合成に関与しているとは当然ながら断言できない。しかし、現時点では、アルケノンの合成を促進する要因としては低温ストレス以外の情報はなく、関連遺伝子の絞込みは困難である。今後、培養条件をさらに様々に変化させ、アルケノンの合成を直接的に制御する環境条件を見出して行く必要がある。アルケノンの蓄積量は定常期に入った培養後期に上昇することから、培地中の栄養塩の欠乏や、細胞状態の変化が合成の引き金になっている可能性も十分考えられる。そのため、現在、低窒素栄養状態での培養を試みている途中である。他にも、この生物の特徴でもある円石形成も培養後期に顕著になることから、石灰化反応とアルケノン合成の関連性も推測される。よって円石形成を促進するリン欠乏条件や、炭酸水素ナトリウムの過剰添加下での、アルケノン合成量の変化から、アルケノン合成の制御要因を特定することも試みるつもりである。
2.RACE法によるEhKasα、EhEloα、EhEloγの全長決定とヘテロな発現系の確立 ESTでは配列情報が3’側に限られており、5’側は途中で切断されていて全長配列を得られない事が多い。EhKasα、EhEloαおよびEhEloγの場合も3’側から数百bp以上の5’側の配列情報は不明である。そこでRACE法を用いて5’側の配列決定を行う予定である。そして全長を取得し、酵母などで発現させ、合成される脂質の中間代謝産物の解析から、当該酵素の特性を明らかにする予定である。
©2007 筑波大学生物学類
|