![]()
![]()
![]()
![]()
![]()
光合成真核生物の細胞内にはエネルギー変換にかかわる2つの小器官があります。いうまでもなく,ミトコンドリアと葉緑体です。これら二つのオルガネラはいずれも独自のDNAとタンパク質合成系をもち,リボソームが70S(Sは沈降係数,細胞質のリボソームは80S)で,さらに抗生物質感受性が原核生物に類似するなどの性質を共有しています。この事実は,この2つの細胞小器官が原核生物の共生によって獲得されたことを示しています。
葉緑体は藍藻に近縁の原核藻類の共生によって生じたことがほぼ確かめられています。そのさらに古い祖先は光合成色素の組成やそのしくみからみておそらく光合成細菌であると考えられます。おそらく相同な電子伝達系が存在することから,ミトコンドリアもまた葉緑体と同じく紅色細菌に近縁である可能性があります。
![]() 16SrDNAの系統樹(葉緑体が単系統であることを示している)
16SrDNAの系統樹(葉緑体が単系統であることを示している)
核の起源は現在でも不明な点が多く,定説はありません。細胞学や分子系統から核は古細菌が真正細菌に共生して生じたという見解が提出されています。このことの真偽のほどはまだ明らかではありませんが,魅力ある仮説といえます。
細胞共生は生物進化に想像以上に深くかかわっており,特に真核光合成生物の多様性実現の原動力となってきたといえます。クリプト藻がよい例です。

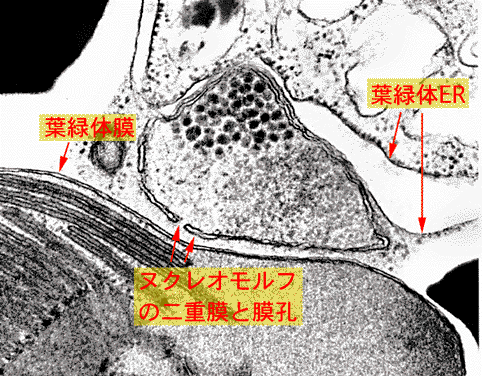
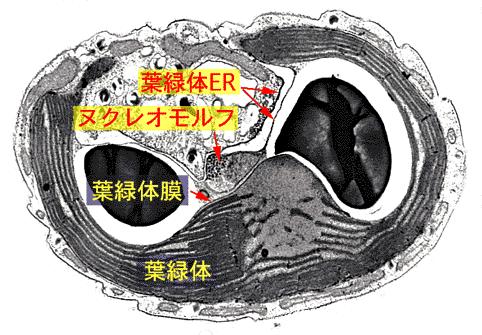 クリプト藻では葉緑体が葉緑体小胞体とよばれる2枚の膜に包まれ,両者の間に2重膜にかこまれ,DNAとRNAをもつヌクレオモルフとよばれる構造があります。リボソームRNAの系統からヌクレオモルフは紅藻類の核に近縁であり,クリプト藻の核とはかけ離れていることが明らかにされました。これは紅藻類に近縁の真核生物が他の真核細胞に共生し,ミトコンドリアを失い,また核が退化してヌクレオモルフに変化したことを示しています。つまりクリプト藻の葉緑体は真核生物の共生によって獲得されたものです。
クリプト藻では葉緑体が葉緑体小胞体とよばれる2枚の膜に包まれ,両者の間に2重膜にかこまれ,DNAとRNAをもつヌクレオモルフとよばれる構造があります。リボソームRNAの系統からヌクレオモルフは紅藻類の核に近縁であり,クリプト藻の核とはかけ離れていることが明らかにされました。これは紅藻類に近縁の真核生物が他の真核細胞に共生し,ミトコンドリアを失い,また核が退化してヌクレオモルフに変化したことを示しています。つまりクリプト藻の葉緑体は真核生物の共生によって獲得されたものです。
クリプト藻の他にも真核細胞同士の共生が数多く知られています。クロララクニオン藻にはクリプト藻と同じようにヌクレオモルフが残されています。褐藻類などの黄色植物やユーグレナ(ミドリムシ)類,渦鞭毛藻類の葉緑体も真核藻類が共生し,共生体の核やミトコンドリアが完全に消失したものと考えられています。
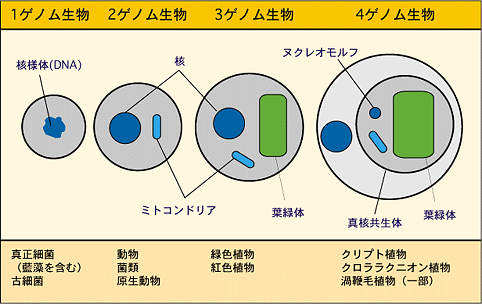 細胞共生がこのように真核生物の多様化に寄与してきたとすると,細胞を構成するゲノム(1種の生物を構成する遺伝子のセット)がいくつの起源をもつかということを生物をみる視点のひとつに加えることができます。原核生物は共生という現象を体験しておらず,したがって1ゲノム生物とよぶことができます(ここでは核の共生起源を想定していません)。ミトコンドリアを獲得した動物や菌類などの生物は2つの生物に由来するゲノムをもつので2ゲノム生物,さらに葉緑体を獲得した緑色植物や多くの藻類は3ゲノム生物ということになります。そしてクリプト藻やクロララクニオン植物は4ゲノム生物です(実際には共生した真核藻類のミトコンドリアがあったはずなので,5ゲノム生物とよぶのが正しいのですが,その証拠はみつかっていないのでここでは数に入れていません)。渦鞭毛植物,ユーグレナ植物,黄色植物なども真核藻類の共生によって葉緑体を獲得したと考えられるので4ゲノム生物に分類されるべきものです。このように真核細胞は異なる履歴をもつ生物のキメラとしてとらえることができます。真核生物が真に理解されるためにはこのような細胞共生によるオルガネラ獲得のメカニズムが理解される必要があります。
細胞共生がこのように真核生物の多様化に寄与してきたとすると,細胞を構成するゲノム(1種の生物を構成する遺伝子のセット)がいくつの起源をもつかということを生物をみる視点のひとつに加えることができます。原核生物は共生という現象を体験しておらず,したがって1ゲノム生物とよぶことができます(ここでは核の共生起源を想定していません)。ミトコンドリアを獲得した動物や菌類などの生物は2つの生物に由来するゲノムをもつので2ゲノム生物,さらに葉緑体を獲得した緑色植物や多くの藻類は3ゲノム生物ということになります。そしてクリプト藻やクロララクニオン植物は4ゲノム生物です(実際には共生した真核藻類のミトコンドリアがあったはずなので,5ゲノム生物とよぶのが正しいのですが,その証拠はみつかっていないのでここでは数に入れていません)。渦鞭毛植物,ユーグレナ植物,黄色植物なども真核藻類の共生によって葉緑体を獲得したと考えられるので4ゲノム生物に分類されるべきものです。このように真核細胞は異なる履歴をもつ生物のキメラとしてとらえることができます。真核生物が真に理解されるためにはこのような細胞共生によるオルガネラ獲得のメカニズムが理解される必要があります。
![]()
![]() 真核藻類の共生に由来する葉緑体をもつ藻類がさらに他の生物に共生する例が知られています。渦鞭毛藻のなかにはクリプト藻を共生体としてもっているものがあります。この場合,クリプト藻はすでに複数の共生を繰り返してきた4ゲノム生物であるので,渦鞭毛藻の核とミトコンドリアを加えて合計6つのゲノムからなっていることになります。
真核藻類の共生に由来する葉緑体をもつ藻類がさらに他の生物に共生する例が知られています。渦鞭毛藻のなかにはクリプト藻を共生体としてもっているものがあります。この場合,クリプト藻はすでに複数の共生を繰り返してきた4ゲノム生物であるので,渦鞭毛藻の核とミトコンドリアを加えて合計6つのゲノムからなっていることになります。
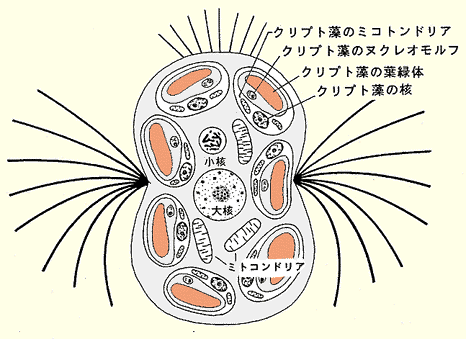
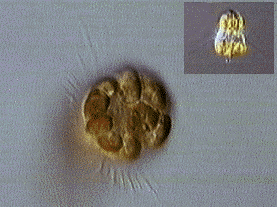 同じように,赤潮をつくる繊毛虫であるメソディニウム(Mesodinium)には多数のクリプト藻が細胞内共生している。これはクリプト藻が起源の異なる少なくとも4つのゲノムをもつことを考えあわせると,繊毛虫の核(ここでは大核と小核で1つの核と数える)を加えて6つの異なるゲノムで1個の種が構成されている。
同じように,赤潮をつくる繊毛虫であるメソディニウム(Mesodinium)には多数のクリプト藻が細胞内共生している。これはクリプト藻が起源の異なる少なくとも4つのゲノムをもつことを考えあわせると,繊毛虫の核(ここでは大核と小核で1つの核と数える)を加えて6つの異なるゲノムで1個の種が構成されている。
![]()
![]()
![]()
![]()
![]()