【導入及び目的】
トロポニンTは筋収縮調節蛋白質トロポニン複合体を構成するのサブユニットのひとつであり、等電点や分子量の異なる多くのアイソフォームをもつタンパク質である。それらのアイソフォームは速筋型、遅筋型、心筋型という3種類の遺伝子に由来する転写産物の選択的スプライシング、また、翻訳後のタンパク質の修飾によって生じ、発生段階特異的、筋組織特異的に発現することが明らかになっている。従って、各遺伝子の一次構造を決定することにより、各アイソフォームの特異的な発現機構の解明への手がかりが得られるものと思われる。
ニワトリ速筋型、心筋型トロポニンTに関しては、 cDNA の塩基配列のみならず、遺伝子の構造がほぼ明らかにされ、複数の異なるアイソフォームを生み出す選択的スプライシングの様相が解明されている。一方、遅筋型トロポニンTについては、タンパク質レベルで筋組織特異的な5 種のアイソフォームの発現が確認され、 7 種類の
cDNA が単離されているものの、遺伝子の構造は殆ど研究されていない。遅筋型トロポニンT遺伝子の転写産物を詳細に調べた結果、5'
側の翻訳開始点から 91 ~ 93 番目にアラニンのコドンを含み、214 番目に C 、218 番目に A を持つ転写産物と、アラニンのコドンを含まず、214
番目に T 、218 番目に G を持つ転写産物が存在することが示された。また、 3' 側では塩基置換が 4 ヶ所で見られた。5'
側のアラニンのコドンの有無や2カ所の塩基置換、また、3' 側の4カ所の塩基置換が選択的スプライシングによるのか、個体差を示しているのか、転写産物の解析から明らかにするのは困難である。そこで本研究では、遅筋型トロポニンT遺伝子の構造を決定し、その選択的スプライシングの様相を解明すること、さらにこの遺伝子の転写調節領域の解析を行って、その発現調節機構を検討することを目的とした。
【方法】
ニワトリの肝臓から抽出した DNA を鋳型とし、 cDNA の塩基配列よりデザインしたプライマーセットを用いて genomic
PCR を行い、ニワトリ遅筋型トロポニンT遺伝子の構造領域を単離した。またこれを pBluescript II KS+ を用いてクローニングし、オートシークエンサーを使用して塩基配列の決定を行った。
【結果及び考察】
genomic PCR を行った結果、ニワトリ遅筋型トロポニンT遺伝子の構造領域は約 3 kb であることが判明した。これはニワトリ心筋型トロポニンT遺伝子の大きさの
1/3 、ニワトリ速筋型トロポニンT遺伝子(全長約 35 kb )のイントロン1とほぼ同じサイズという非常に小さいものであった。遅筋型トロポニンTは速筋型及び心筋型トロポニンTとその
cDNA の大きさに関してはほとんど差がないため、この遺伝子のイントロンのサイズは非常に小さいと考えられる。また、塩基配列の決定の結果、5'
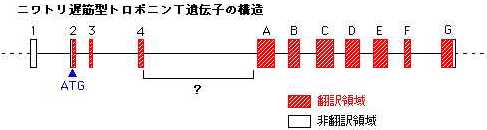
側の エクソン 1 から 4 まで、 3' 側の 7 つのエクソン(エクソン A 、B 、C 、D 、E 、F 、G )の構造が明らかになった。エクソン
1 と 2 、エクソン G は各々 5' 非翻訳領域と 3' 非翻訳領域を含んでいた。
先に述べたアラニンのコドン GCA は、最初の G がエクソン 4 の一番最後に、残りの CA はエクソン 5 に含まれていた。イントロン
4 の塩基配列を決定することにより、アラニンのコドンの有無が選択的スプライシングによるものか否かを明確にすることができると思われる。残されたエクソン
4 とエクソン A の間の約 1 kb には GC が非常に多く含まれ、塩基配列の決定が困難であるが、現在解析が進んでいる。また、今後は構造領域の全塩基配列の決定を完了させるのに加え、転写調節領域についても塩基配列の決定・解析を行い、ニワトリ遅筋型トロポニンT遺伝子の発現調節・選択的スプライシングの調節機構を明らかにしていきたい。