Pyrococcus horikoshiiOT3株の推定リジン生合成クラスターのBacillus subtilisへの導入による機能解析
赤津 裕一 (筑波大学 生物学類 4年) 指導教官:星野 貴行 (筑波大学 生物科学系)
<導入・目的>
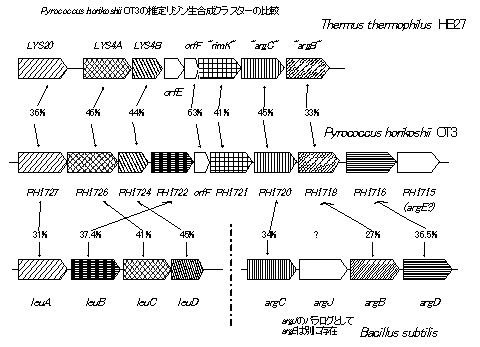 当研究室では高度好熱性真正細菌Thermus
thermophilus HB27のLys生合成が、バクテリアで一般的なジアミノピメリン酸経路(DAP経路)ではなく、カビ・真菌類でのみ報告されていたa-アミノアジピン酸を中間体とする経路によって行われていることを発見した。また、T.
thermophilus のLys生合成クラスターと高い相同性を示す領域が超好熱性嫌気性古細菌Pyrococcus.horikoshiiOT3ゲノム中にも見出され、同菌株も同様の経路によりLys生合成を行っていることが推定された。同時にP.
horikoshiiの遺伝子クラスターは、上流4遺伝子(PH1727-PH1722)が他の生物のLeu生合成遺伝子と、下流の3遺伝子(PH1720-PH1716)がArg生合成遺伝子と有意な相同性を示している。P. horikoshii OT3はLys、Arg、Leuに対して要求性を示さず、またそのゲノム上にはこの領域以外にLeu、Arg生合成遺伝子、あるいはジアミノピメリン酸経路を構成するLys生合成遺伝子と相同性を示すような遺伝子は存在していない。以上から、この領域はP.
horikoshii OT3中でLys、Leu、Argの3種のアミノ酸生合成に関与することが推察された。
当研究室では高度好熱性真正細菌Thermus
thermophilus HB27のLys生合成が、バクテリアで一般的なジアミノピメリン酸経路(DAP経路)ではなく、カビ・真菌類でのみ報告されていたa-アミノアジピン酸を中間体とする経路によって行われていることを発見した。また、T.
thermophilus のLys生合成クラスターと高い相同性を示す領域が超好熱性嫌気性古細菌Pyrococcus.horikoshiiOT3ゲノム中にも見出され、同菌株も同様の経路によりLys生合成を行っていることが推定された。同時にP.
horikoshiiの遺伝子クラスターは、上流4遺伝子(PH1727-PH1722)が他の生物のLeu生合成遺伝子と、下流の3遺伝子(PH1720-PH1716)がArg生合成遺伝子と有意な相同性を示している。P. horikoshii OT3はLys、Arg、Leuに対して要求性を示さず、またそのゲノム上にはこの領域以外にLeu、Arg生合成遺伝子、あるいはジアミノピメリン酸経路を構成するLys生合成遺伝子と相同性を示すような遺伝子は存在していない。以上から、この領域はP.
horikoshii OT3中でLys、Leu、Argの3種のアミノ酸生合成に関与することが推察された。
本研究では遺伝子操作の容易な枯草菌を宿主として発現させることにより、P.
horikoshii OT3のこの遺伝子領域がLys、Leu、Arg生合成に関与しているかどうかを明らかにすることを目的とする。
<方法>
Leu、Arg生合成に関与する遺伝子を個別に破壊した枯草菌菌株を新たに作製し、それぞれの破壊株にP.
horikoshii OT3の推定Lys生合成クラスター中の対応遺伝子を導入して、変異の相補(Leu、Argに対する要求性の回復)により導入遺伝子がそれぞれLeu、Arg生合成に関与するのかどうかを明らかにする。Lys生合成に関しては、枯草菌ではDAP経路を利用しているので、最終段階の反応を司る遺伝子(lysA)の破壊株を作製し、この株に対してP. horikoshii OT3の推定Lys生合成クラスター全長を導入する。
第一段階としてLeu生合成遺伝子に着目し、枯草菌のleuオペロンを構成する各遺伝子を、各ORF内部を大きく欠失させ、それを枯草菌の相当遺伝子と置換することにより破壊する。次にP.
horikoshii OT3の対応遺伝子のORFをPCR増幅し、そのC末端にFlag-tagを挿入する。この構築を枯草菌xyl promoterの下流に結合して各変異株ゲノムのamyE locusに挿入する。このようにして作製した菌株についてLeuに対する要求性を、枯草菌の生育上限温度である52~55℃(培地組成、塩濃度によって可変)付近での最少培地での生育により検討し、P.
horikoshii
OT3の導入遺伝子がLeu生合成に関与しているかどうかを明らかにする。なお導入遺伝子の発現の有無については、Flag-tagに対する抗体を用いたWestern
blottingにより解析する。
<結果・考察>
現在、枯草菌のleuA、leuB破壊株の構築に成功し、leuC、leuD破壊株の構築を行っている。導入するP. horikoshii OT3遺伝子については、各ORFのPCR増幅及びクローン化は完了し、現在Flag-tagの付加を行っている。またamyE locusに挿入するためのプラスミドについても構築を行った。
P. horikoshii OT3は超好熱性古細菌なので、遺伝子産物が常温で活性を持たないことがある。実際に大腸菌で発現させた場合には、産物が活性を持たなかったという報告もある。それに対して枯草菌は生育上限温度が大腸菌より高く、GC含量も同程度なので、P. horikoshii OT3遺伝子を発現させる際のより良い宿主として利用可能であると考えられる。