梶原 誠 (筑波大学 生物学類 4年) 指導教官:田仲 可昌 (筑波大学 生物科学系)
<導入・目的>
転写因子は遺伝子の発現を制御する上で重要な役割を担っている調節因子であり、遺伝子の時間的・空間的制御を必須とする多細胞生物の発生において中心的な役割を果たしている。しかしながら、発生・細胞分化に関わる遺伝子発現がいつ・どのような転写因子によって調節されているのかは不明な点が多い。
本研究室では細胞性粘菌Dictyostelium discoideumの発生をモデルとした多細胞体形成や分化機構に関する研究が、細胞性粘菌のゲノム解析によって得られた転写因子候補遺伝子に注目し進行している。すでにいくつか転写因子候補遺伝子の破壊株が作成され、転写因子の多細胞体形成における遺伝子調節の解析が始まろうとしている。
その研究の一環として、得られた破壊株の発生過程・細胞分化パターンを詳細に観察するために、細胞性粘菌の分化パターン特異的に発現する既存のマーカー遺伝子コンストラクトを破壊株に導入することを試みた。しかし、このマーカー遺伝子コンストラクトはゲノムにランダムに組み込まれるタイプのため、組み込みによる二次的変異を引き起こす恐れがあり、さらに形質転換効率が非常に低く今後の研究に適切でないと判断した。そこで本研究では、ゲノム組み込み型ではないマーカー遺伝子コンストラクトの作製を行った。
<材料・方法>
既存のマーカー遺伝子コンストラクトはゲノムにランダムに組み込まれるタイプであり、検出方法はβ‐galactosidase活性による方法であったが、新しく作製するマーカー遺伝子コンストラクトはゲノムに組み込まれず染色体外で発現するextrachromosomal型で、活性の検出はGFPの蛍光発色による方法にした。以下にその作製手順を示す。
1)既存のゲノム組み込み型のマーカー遺伝子コンストラクトから、粘菌の分化パターン特異的(時間的・空間的)に発現する遺伝子(ecmA,ecmB,ecmO,ecmAO,pspA)のプロモーター部分を切り出す。
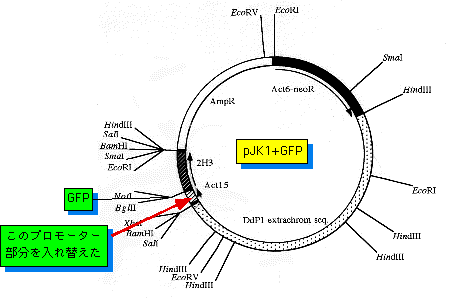
2)新しくベースとなるextrachromosomal型のGFPベクター(右下図参照)から、Act15プロモーター部分を抜き出す。
3)DNA断片をつなぎ合わせる。
4)それぞれのプロモーター下流で、GFPのORFのフレームが合っていることを、塩基配列を決定して確認する。
<結果>
作製したプラスミドベクターは右図のようになる。今後の予定としては、
挿入したプロモーター部分が設計した位置と同じ向きに挿入されて
いるかどうかを塩基配列を決定して確認する。その後、これらの
マーカー遺伝子コンストラクトを野生株の粘菌細胞に導入して
GFPが分化パターン特異的に発色するかを調べる。