|
つくば生物ジャーナル Tsukuba Journal of Biology (2004) 3: TJB200406MY.
活性酸素は毒ですか?
植物におけるストレスの存在意義
柳田 元継(岡山県生物科学総合研究所(RIBS)細胞工学部門)
はじめに
「活性酸素は生物にとって毒である。」これはまぎれもない事実であり、いまや一般の人にまで知られている。が、「活性酸素は毒でしかないのか?」と考える人は多くない。私がこの発想にいたるまで、そしてその新たな答えを見つけ出すまでの過程を本稿で綴っていくことにする。
活性酸素との出会い
酸素について「生物の呼吸で使われ重要なもの」、よくて「多くの代謝反応に使われる基質」程度の幼稚かつ一面的な知識しかもっていなかった私に衝撃を与えたのは、生物学類3年時の松本先生の植物制御学の授業であった。除草剤のひとつにクロロフィル合成を阻害するタイプのものがある。植物に処理されたこの薬剤はクロロフィル合成が行われる葉緑体に移行し、合成酵素のひとつのプロトポルフィリノージェンオキシダーゼを阻害し、前駆体のプロトポルフィリノージェンを大量に蓄積させる。この前駆体はあふれ出す形で細胞質へと移動し、細胞膜付近にあるとされる酸化活性により酸化され、プロトポルフィリンⅨ(ProtoⅨ)に変換される。このProtoⅨは光増感色素であり、光が照射されると活性酸素である一重項酸素を発生させる。この一重項酸素が引き金となり様々な活性酸素分子種(スーパオキシド(O2-)や過酸化水素(H2O2)等)が関与する脂質過酸化の連鎖反応が起こり、白化、しおれ等の症状を呈し枯死する。この活性酸素を発生させるまでの巧妙な仕組みにも感動したが、それ以上に益でもあり、毒でもある酸素の両極端な姿に興味をそそられずにはいられなかった。この後、松本先生のラボで研究を行うことになったが、大きなテーマとして上記除草剤の選択性のメカニズムに取り組んだ。注目したのは各々の植物が持つ抗酸化能力であった。
この酸素傷害を防御する能力はすべての生物が持っていると考えてよい。現在より1/10000の酸素濃度であった30億年以上前に出現したとされるほとんどの嫌気性細菌にも活性酸素のひとつのO2-を消去するスーパーオキシドドディスムターゼ(SOD)が見出されている。藍藻をはじめとする光合成生物により酸素が大気に供給されはじめたが、当初、酸素は他の生物にとって毒ガス以外の何ものでもなかったと思われるが、この中で酸素傷害を回避する能力を持っていた生物のみが、酸素を利用する能力を獲得したと考えられる。
さて、この抗酸化能力と除草剤選択性との関係を明らかにするために、SODのような抗酸化酵素の活性やグルタチオン(GSH)のような抗酸化物質の含量を調べた。その結果、薬剤処理に対してより枯れにくい植物種ほどこれらの値は高く、一方、枯れやすい植物種においては低かった。以上より活性酸素を発生させるタイプの除草剤の選択性をきめる要因の一つとして、植物が元来有する抗酸化能力が深く関わることが明らかになった(Yanagida et al. 2001)。よって、活性酸素は生物にとって明らかにストレスでその命を脅かすもの、逆に抗酸化能力はその毒性を減らすことによる生命維持機構であるという構図が私自身に刷り込まれた。
活性酸素は毒だけなのか?
2000年4月から岡山県生物科学総合研究所に勤務することになった。そこで今のボスでもある小川健一博士と出会う。「活性酸素は植物の成長を制御する」ことをデータ(Ogawa and Iwabuchi 2001)とともに説明を受け、研究テーマについてのディスカッションが続いた。その中で活性酸素による成長促進効果については、活性酸素の毒性についてみてきた私には信じがたいものがあった。が、大学時代に一つだけこれを支持するデータを得ていたことを思い出した。除草剤の濃度に依存し生育は抑制されるのだが、植物が枯れないごく薄い濃度では無処理区に比べてむしろ生育がよくなっていたのだ。活性酸素は「毒である」から「毒だけなのか?」、そして「生物に必要なものではないか??」と徐々にだが、活性酸素を異なる角度で見始めるようになる。
活性酸素の新たな解釈
我々は光が強くなれば早く花が咲くことや、寒い冬を乗り越えて春を迎えて花をつけることをこれまでに経験的に知っている。環境ストレスが誘導する花成である。環境ストレスはその種類が違っても、結果、活性酸素を発生し植物に酸化ストレスを与える。例えば低温ストレスの場合、カルビンサイクルにおける酵素反応が抑えられ、光化学系で作られたNADPHが蓄積し、NADP+が欠乏する。しかし、光エネルギーから変換された電子の流れは止まらないため、過剰な電子が酸素に渡りO2-が発生する。こうした低温ストレスに対する防御反応としてGSH量が変動することが報告されている。
GSHはユビキタスに存在するトリペプチドであり、γ-グルタミルシステインシンテターゼ(γ-ECS)とグルタチオンシンテターゼによる連続した2段階の反応で合成される。GSHは植物においていくつもの機能を有している。抗酸化能力のほか、生体異物の解毒においてGSH抱合反応の基質となっている。また重金属をトラップするファイトケラチンの前駆体でもある。さらに、GSHが細胞分裂や根毛成長、管状要素分化、アントシアニン蓄積、糖代謝の制御等にかかわることが報告されている。
とりわけ小川博士はシロイヌナズナの変異体を用いて、GSHが花成の制御にかかわることを示した(Ogawa et al. 2001)。しかしGSHの正確な役割については不明のままであった。それがゆえに、低温ストレスが誘導する花成にGSHがエッセンシャルであるかどうか評価することは意義深いことであった。そこで抽だい(茎伸張)に低温を絶対的に必要とするロゼット植物のトルコギキョウを用いてこの問題に取り組んだ。トルコギキョウは抽だい後、花芽を形成するので、栄養成長から生殖成長への相転換を抽だいで評価した。
まず、トルコギキョウの抽だいを誘導するのにGSHが低温に取って代わることができるかどうか調べたところ、低温処理をしていないにもかかわらず、GSHにより抽だいが促進された。このとき内生のGSH量が上昇していたことから、GSHが抽だいのinducerと考えられた。そこで次に低温による抽だいがGSHに起因することを示すために、低温処理と同時にGSHの合成阻害剤を処理した。阻害剤は抽だいを完全に抑制し、低温による抽だい促進効果を完全に打ち消した。さらに、この阻害剤による抑制はGSHの同時処理により、GSH濃度依存的に回復した。以上から低温による抽だい誘導過程にGSH合成が必要なことがわかった。
低温処理は4週間、15℃で行われたが、このとき植物の葉は薄い緑色を呈し、実際クロロフィル含量も低下しており、典型的な低温傷害の症状を示した。そこで低温処理中のGSH合成と酸化ストレスとの関係を調べるために、酸化ストレスによる脂質の過酸化の指標であるthiobarbituric
acid reactive substances(TBARS)の経時的変化を調べた。この結果から、処理開始1週間で最も酸化ストレスを受けていたことが示された。またGSH含量も低温処理1週間高いレベルを示し、TBARSの変動のプロファイルと一致した。さらにGSH量が上昇した期間のGSH合成を詳細に調べたところ、前駆体であるシステインやγ-グルタミルシステインの変動は見られなかった。またGSH合成酵素のγ-ECSの活性はこの間上昇したが、γ-ECSをコードするGSH1のmRNA量は変化していなかった。シロイヌナズナにおいてストレス誘導性のγ-ECSの転写後活性化が報告されているので、こうした結果はγ-ECS活性の上昇が低温による酸化ストレスにより引き起こされ、これがGSH合成促進のキーステップであること反映している。よって、図に示したようにトルコギキョウにおける低温ストレス誘導性の抽だい機構は、低温によりGSH合成酵素のγ-ECSが活性化しGSHが合成され、このGSHが抽だいを促進することが示された(Yanagida
et al. 2004)。
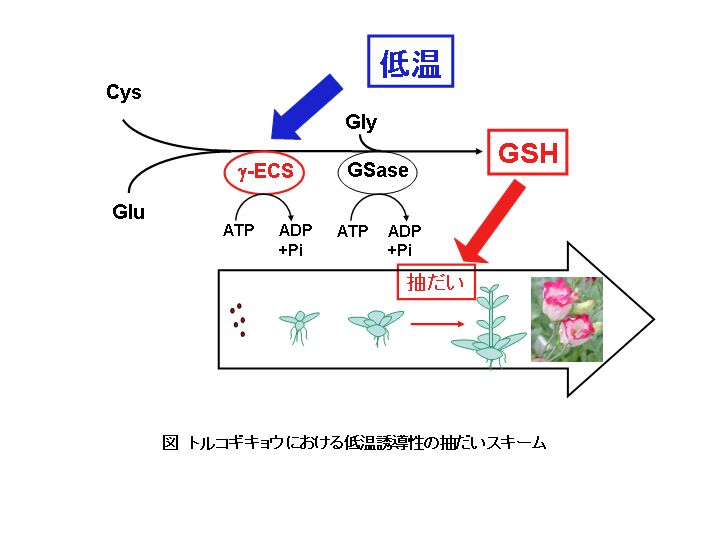
以上のことから「活性酸素は毒」ではなく、トルコギキョウの抽だいのために必要なGSH合成を活性化するための「必要な刺激」であることがわかる。またGSHに関してはこれまで、活性酸素を消去して生物の寿命を延ばすとされていたが、実際には花成を促進し、結果として植物のライフサイクルを短くする方向に働くことがわかった。未発表のデータではあるがグルタチオンの合成の抑制はストレスによる植物の老化を遅延させ、個体の死も抑制することもわかってきた。
最後に
物事を多角的に理解することがきわめて重要であることは理解していても、それを研究に、あるいは普段の生活において実行することは難しい。ヒトはとかく困難から逃げ出したがる。しかし、子どもから大人に成長するには、また大人が健全な人生を送っていくにはあえて困難に立ち向かわなければならないときがある。最近ストレスという言葉を盛んに耳にするが、想像してみてほしい。ストレスがすべて取り去られた状態は、われわれに幸福をもたらすであろうか?
謝辞 本研究を支えて下さった多くの先生方、また研究室の皆様に感謝いたします。特に、今の研究所に来て以来、首尾一貫して面倒を見ていただいた小川博士、研究を始めてから今日まで温かく見守って下さった松本先生に深く感謝いたします。
参考文献
Yanagida, M., Matsumoto, H. and Usui, K. (1999) J. Weed Sci. and Tech.
44(1): 67-76
Ogawa, K. and Iwabuchi, M. (2001) Plant Cell Physiol. 42: 286-291.
Ogawa, K., Tasaka, Y., Mino. M., Tanaka, Y. and Iwabuchi, M. (2001) Plant Cell Physiol. 42: 524-530.
Yanagida, M., Mino. M., Iwabuchi, M. and Ogawa, K. (2004) Plant Cell Physiol. 45: 129-137.
Communicated by Hiroshi Matsumoto, Received July 9, 2004.
©2004 筑波大学生物学類
|