|
つくば生物ジャーナル Tsukuba Journal of Biology (2006) 5: TJB200610TM.
動物系統分類学 ―私の昔のテキストから―(連載第3回)
牧岡 俊樹(元 筑波大学 生物科学系)
第2章 生物の中の動物
第1項 生物の世界の全体像
近年の電子顕微鏡や分子生物学の手法による研究の結果、地球上の生物の体は構造と機能の著しく異なる2種類の細胞、つまり原核細胞 (prokaryotic
cell) または真核細胞 (eukaryotic c.) のどちらかからできていることがわかった。ゆえに生物はまず第一に、原核細胞からできている原核生物
(Prokaryota) と真核細胞からできている真核生物 (Eukaryota) とに分けられるべきである。この区分は従来の最大の分類群である界を超えた新しい区分であり、まだ公式の名称ではないが、ドメイン(Domain、超界)という名がほぼ定着している。最近は原核生物をさらに2分して古細菌ドメイン(Domain
Archaea)および細菌ドメイン(Domain Bacteria)とし、真核生物ドメイン(Domain Eukarya)と合わせて生物界を3つのドメインとすることが多い(図4)。
しかしこれら3つのドメインの間でも、遺伝子の構成分子、タンパク質合成の方法、遺伝暗号の読み方など、生命の最も本質的な部分は共通で、もとは同じ祖先から生じたものであることがわかる。
1─1 古細菌類と細菌類
原核細胞からできている生物で、多くは単細胞生物であるが、細菌類には群体 (colony) や分化した原核細胞から成る原核多細胞体を作るものもある。減数分裂と接合(受精)を伴う有性生殖を行なわないので、生物学的種の概念や生殖的隔離による種分化の概念は当てはめられない。また細胞の構造が簡単で表現型上の特徴が少なく、今までに命名された種(形態種)の数も上種分類群の数も少ない。ゆえに従来はただ1つの界(モネラ界
Kingdom Monera)にまとめられることが多かったが、生命の起源以来の永い歴史から見ても、多数の進化の方向を含む多様な群であり、多数の界に分けられる筈である。
近年はゲノムの塩基配列の解読が進んで、遺伝子型の特徴を考慮した分類法が研究されており、また土壌や水中など通常の環境から採集された DNA 断片から、多数の未発見の原核生物の存在が推定されるようになった。今までに知られている古細菌類は無酸素、高温、高塩分濃度、低
pH など、他の生物が棲めない極端な環境に棲むので、原核生物の中での遺存群(古い型の生物の生き残り:生きた化石)とみなされていたが、通常の環境中の
DNA 断片には古細菌類の特徴を示すものも多く、未発見の多くの古細菌類が通常の環境中に存在するらしいことがわかってきた。古細菌類のゲノムの特徴などには真核生物に似た点があり、真核生物の出発点となった原核生物の候補として、現生の高度好熱好酸古細菌類に近い祖先古細菌類が想定されている。
1─2 真核生物
真核細胞からできている生物で、単細胞のものも多細胞の複雑なものもあり、多様な分類群を含む。多細胞の真核生物を動物界 (Kingdom Animalia)、植物界
(Kingdom Plantae)、菌界 (Kingdom Fungi) に区分することが多く、また単細胞体や分化程度の低い多細胞体など体制の単純な真核生物を集めて原生生物界
(Kingdom Protista) を設けることもある。原生生物界は他の界のように特定の進化の方向によってではなく、体制の発達の程度によって区分された人為群である。近年、主にゲノムのデータにより、原生生物の中に、界に相当する大きな群として、ユーグレノゾア類(Euglenozoa:ミドリムシ類、トリパノソーマ類)、ストラメノパイル類(Stramenopila:褐藻類、珪藻類、卵菌類など)、アルべオラータ類(Alveolata:渦鞭毛虫類、繊毛虫類、アピコンプレクサ類)
などをまとめる傾向があり(第3章)、原生生物は界を超えた群(上界 Superkingdom)として再編成されつつある。
|
コラム10:膜進化説と共生説
真核細胞は原核細胞よりも大きく、小胞体やゴルジ体や核膜、またミトコンドリアや葉緑体などの細胞小器官をもつ。DNAは遺伝子部分(構造配列、エクソン)の他に多くの介在配列(イントロン:タンパク質合成に使用されない塩基配列)を含んで長く、複数の染色体に分かれて核中にある。タンパク質合成の場であるリボソームも原核細胞のもの(70S)よりやや大きい(80S)。酸素呼吸の場であるミトコンドリアと光合成の場である葉緑体はともに外膜と内膜の2枚の膜から成り、独自の環状DNAをもつ。
真核細胞は原核細胞から進化したと考えられるが、そのでき方には2つの考え方があった。膜進化説では、真核細胞の祖先となった原核細胞の細胞膜が細胞内に深く陥入し、複雑に分岐し、折りたたまれて小胞体やゴルジ体や核膜などを形成したと考える。また共生説では、真核生物の祖先となった原核生物の細胞内に他の原核生物(好気性細菌や藍藻)が共生してミトコンドリアや葉緑体になったと考える。
両説は相補的な関係にあり、真核細胞の起源は両説を併せて説明するのがよいと思われる。しかしそれでも、原核細胞の環状のDNAが真核細胞では染色体ごとに分断されていることの由来や、介在配列の起源、また転写(DNAからmRNAが作られる)された介在配列部分を切り落とす(スプライシング)機能の起源などまだ不明の点も多い。
|
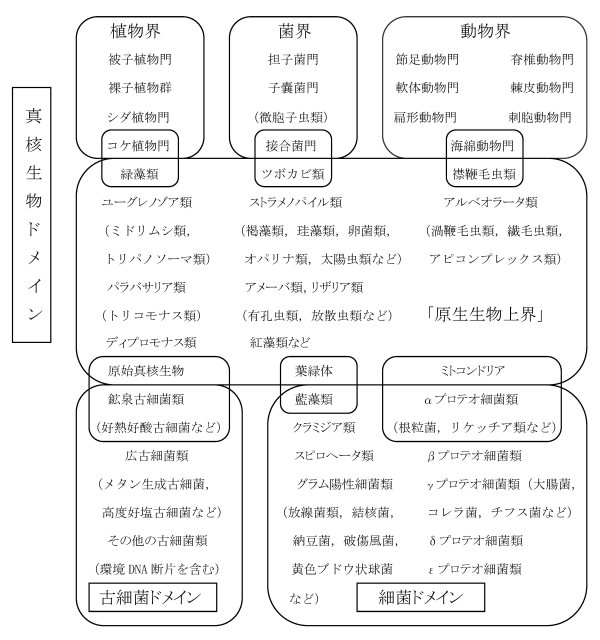
図4 地球上の生物の大分類と類縁
真核生物(ユーカリア)の基本群である原生生物は、細胞本体は古細菌類(アーケア)のうち、おそらく現在の好熱好酸古細菌類に近い祖先から生じ、その後ミトコンドリアは細菌類(バクテリア)のうちのαプロテオ細菌類に近い酸素呼吸をする祖先が、また葉緑体は光合成細菌である藍藻(らんそう)類の祖先がそれぞれ細胞内共生することによって生じたとする見解(共生進化説)による。真核生物の植物、菌類、動物はそれぞれ独自の原生生物を祖先として生じた多細胞体の界である。原生生物は多様な方向に進化した大きな群で、多数の界に分けられるべきであるが、その分け方にはまだ議論が多いので、ここでは仮に原生生物上界とし、その中の代表的な類を示しておく。原生動物との関係については表2を参照。古細菌類と細菌類も同様に多くの界を含む筈であるが、ここではドメイン内の主な類を示すにとどめておく。
第2項 生物の界区分と動物界
2─1 界区分とその変遷
動物と植物の2大群の区分は人類の歴史とともにあったと思われるが、リンネ(Linne, 1735) の分類体系では最大の分類群として動物界および植物界が設けられた(2界説)。ヘッケル(1866)はさらに原生生物界を設けて3界とし(図2)、また後には植物界から菌界が分けられるなど界の数は増えたが、原核生物と真核生物が区別されたのは電子顕微鏡が普及した20世紀中頃以後のことである。最近までは、モネラ
(原核生物) 界を含むホイッタカー(Whittaker, 1969) やマーギュリス(Margulis, 1974) らの5界区分が広く用いられていたが、人為群であるモネラ界や原生生物界の再編成が進むとともに、界の数はさらに増える傾向にある。実際の生物の歴史では、原核生物、真核生物のそれぞれで、界に相当する多くの方向への進化がくりかえされたに相違なく、表現型と遺伝子型の両面の解析から、それらの多様な進化の方向の解明が進められている。
|
コラム11:ゲノムの水平移動と系統
親から子へ、また元の種から新種へゲノムが伝えられる通常の垂直移動に対して、類縁の遠い生物間でウイルスのはたらきなどによってゲノムの一部が伝えられることがあり、これをゲノムの水平移動という。また細胞内共生によってある生物の細胞全体が別の生物の細胞内に入って一体化することがあり、多くの場合、共生者のゲノムの一部が宿主のゲノムに移行(水平移動)する。
単細胞生物では、摂食や光合成などの生活活動をしている個体がそのまま無性生殖あるいは有性生殖(真核生物の場合)によって増殖するので、ある生物の1個体に別の生物から水平移動によって伝えられたゲノムの一部は、受け入れた生物のゲノムに組み込まれ、それが表現された結果がその生物の生存や生殖に不利な影響を与えなければそのまま残り、垂直移動によって子孫に伝えられ、別の生物の表現形がその生物に定着する。たとえば病原性大腸菌
O-157の毒素遺伝子は赤痢菌から水平移動したものと考えられている。またバイオテクノロジーでは、この現象を利用して大腸菌にヒトのインスリンや成長ホルモンの遺伝子を組み込み、これらのホルモンを大量に作らせることができる。
多細胞生物では、体細胞系列に水平移動したゲノムは1世代限りで垂直移動はしないが、生殖細胞系列に水平移動したゲノムは宿主のゲノムに組み込まれ、その結果が宿主の生存や生殖に不利な影響を与えなければ、そのまま垂直移動によって子孫に伝えられる。たとえばショウジョウバエのP因子と呼ばれるトランスポゾン(transpozon)は、外部から水平移動によって持ち込まれ、ショウジョウバエのDNAの一部として垂直移動している。現在では、P因子を取り出して任意の遺伝子を結びつけ、遺伝子導入のベクター(vector)としてDNA中に再挿入する技術が確立し、分子遺伝学の重要な研究手段として活用されている。
真核生物のミトコンドリアや葉緑体の共生説による起源はすでに定説化しているが、これは単細胞生物の個体レベルでの水平移動であり、ゆえにゲノムの全部が伝えられる。単細胞の原生生物の間には、近年多くの個体レベルの水平移動の例が知られて来ており、その多くは葉緑体をもたない原生生物に葉緑体をもつ原生生物が共生し、一体となったものである。これは真核生物の祖先における葉緑体(藍藻類)の共生を最初の共生とすれば、二度目の共生ということになる。このような場合、共生体のゲノムの一部が宿主細胞の核ゲノム中に移行し(水平移動)、それ以外は共生体に残っているが、これは両者の共有部分を受け持つ遺伝子と両者の独自の部分を受け持つ遺伝子をそれぞれが分担してもっているのであろう。
別の進化系列で確立した遺伝子型と表現型の一部あるいは全部が一緒になる、つまり2種あるいはそれ以上の生物が水平移動によって1種の生物になるということは、従来の垂直移動による系統の分岐とは別の系統関係であり、一旦垂直的に分岐して離れた枝同士、時にはかなり遠い枝同士がつながって網目状の系統関係を形成することを意味している。水平移動が原理的にはどの生物群でも起こり得るものだとすれば、これまでの系統論は、近年の分岐論も含めて大きな見直しを必要とすることになるだろう。
|
2─2 動物界
動物界に属する生物は従属栄養生物 (heterotrophic organism) で、主として捕食 (predation)により、他の生物を自身の食胞(food
vacuole)内または消化管(digestive tract, alimentary canal)内に取りこみ、消化(digestion)して吸収する。小型の動物や幼生は鞭毛
(flagellum, pl. flagella) や繊毛 (cilium, pl. cilia) の運動によって水中を活発に泳ぐが、大型の動物は筋肉 (muscle) が発達し、強力な運動能力がある。また筋肉の活動の調節や外界からの刺激の感受のために神経系
(nervous system) や感覚器官 (sensory organ) が発達する。
動物界はまとまった界であるが、人為群である原生動物 (Protozoa) の扱い方によってその範囲が異なる。従来の動物界から原生動物をすべて原生生物界に移せば、動物界は多細胞動物(multicellular
animal, 後生動物 Metazoa)だけになり、原生動物中にいると思われる多細胞動物の姉妹群は動物界から外れることになる。しかし、従来のようにすべての原生動物を動物界に入れれば、多細胞動物と系統的な関係のない原生“動物”が動物界に含まれ、動物界は多系統(polyphyletic:共通祖先を想定できない)群になる。本講は前者の立場をとり、原生生物中の襟鞭毛虫類(Choanoflagellata)を動物界の姉妹群と認める。だが伝統的な原生動物の名前と概念はまだ一般に広く残っているので、本講でも参考群としてとり上げておく(第3章)。
2─3 初期の動物界
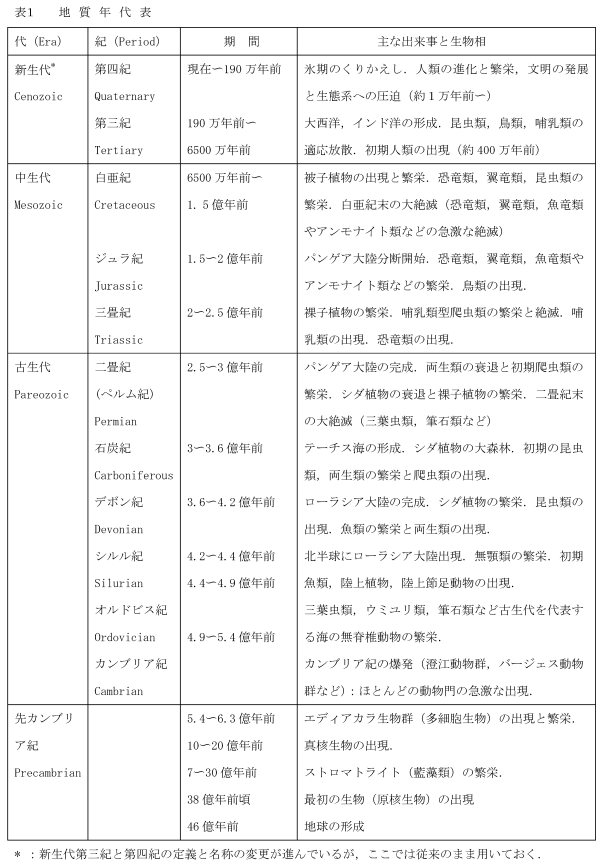
生命の始まりと考えられている約38億年前から、約5.4億年前の古生代カンブリア紀に至る長い先カンブリア紀を通じて、生物の化石は少なく、そのほとんどは原核生物である。
真核生物と思われる比較的大型の単細胞生物の最も古い化石は約20億年前に出ているが、真核生物の特徴である減数分裂(有性生殖)の結果と思われる4細胞の集合体の化石は約10億年前に出ている。多細胞の真核生物の化石としては約6.3-5.4億年前、先カンブリア紀末期のエディアカラ生物群(Ediacara
biota)の印象化石がある。生物体の実体はなく、海底面に圧痕だけが残ったものである。現在の刺胞動物や扁形動物に似たものや、さらに環形動物や節足動物に似た化石もあるが、現在のどの動物門に属するのか、あるいは動物であるのかさえも不明とされる化石もある。
約5.4億年前にはじまる古生代カンブリア紀に入ると、多様な動物化石(澄江動物群、バージェス動物群など)が一斉に出現する。これらの化石はエディアカラ生物群とは著しく違って、印象化石よりも、殻やクチクラなど化石になりやすい硬い部分をもつ実体化石が主であり、しかも古い種類ではあるが現在の動物界の多くの門に入る種を含んでいる。化石の少ない先カンブリア紀に続くこの大量の化石の出現は非常に急激で、これをカンブリア紀の爆発(Cambrian
explosion)と呼ぶことがある。
カンブリア紀の爆発が実際に短期間の多様な進化の結果であるのか、あるいは化石に残っていない先カンブリア紀の進化の結果がカンブリア紀に入ると一挙に化石となって表面化したのかについては多くの議論がある。前者の議論では、短期間に多くの動物門が一斉に出現するような、進化の歴史に例のない急速な進化のしくみや、そのような進化をもたらした原因の説明が必要である。後者の議論では、先カンブリア紀末期には化石として残らなかった多くの動物が、カンブリア紀に入ると急に化石として残るようになった理由や、そのような変化の起こった原因の説明が必要である。
だが生物の進化では、門が突然生じることはなく、最初は必ず新しい種として生じる。その種から次々に新しい種が生じ、やがてそれらの種の間に、属や科や目や綱や門に相当する隔たりが生じてくるにはそれなりの時間が必要である。近年、微小な化石を探索する技術の進歩により、先カンブリア紀末期の微化石の中に環形動物や軟体動物の体の一部と思われる化石や動物の胚や幼生の多様な化石が次々に発見され、エディアカラ生物群の時代の間に、体のやわらかい、あるいは小型の種類を中心に、カンブリア紀の爆発に向かう多細胞動物の進化が進行していた様子が明らかになり始めている。
|
コラム12:雪玉仮説と捕食者仮説
近年の雪玉仮説(snowball earth hypothesis)では、約7億年前、地球は赤道付近も含めて海と陸の全面を厚い氷に覆われ(全球凍結)、当時の生物はほとんど絶滅したが、その後、氷が融けるとともに生き残った生物が急速に増殖し、進化してエディアカラ生物群を生み出し、さらにカンブリア紀の爆発の遠因にもなったと考える。大規模で急激な環境の変化による生物の大絶滅と、生き残った生物が環境の回復とともに急速に増殖し進化することは、たとえば中生代白亜紀末の小惑星の衝突に伴う環境の急変により恐竜類や翼竜類などが絶滅し、環境の回復後の新生代には哺乳類や鳥類が急速に進化した例などにも見られる。約7億年前の全球凍結が事実であれば、それはその直後のエディアカラ生物群の出現と繁栄のきっかけになった可能性がある。だが全球凍結の終わりからカンブリア紀の爆発の始まりまでには約1億年(カンブリア紀の長さの約2倍)という長い時間があり、全球凍結とカンブリア紀の爆発とを直接結びつけるのはやや無理があると思われる。
捕食者仮説(predator hypothesis)では、先カンブリア紀末にもすでに多くの動物門が繁栄していたが、それらはエディアカラ生物群と同じく体の柔らかい種類が多く化石になりにくかった。やがてそれらの中に大型で強力な捕食者(肉食動物)が出現し、体の柔らかい動物は捕食されて次々に絶滅し、各動物門の中で硬い殻やクチクラなどで身を守ることのできた少数の種類が生き残って急速に増殖し、進化したのがカンブリア紀の爆発であると考える。カンブリア紀初期の硬い部分や殻をもった多様な動物の中にもアノマロカリスのような「大型で強力な捕食者」がいたが、先カンブリア紀末期の化石の少ない時代の生態系にもやはりそのような最高位の二次消費者が存在したであろう。だが、それらがたとえば大型のクラゲあるいは肉食性の軟体動物のようなものであったとしても、それらはやはり体が柔らかくて化石になりにくかったと思われる。この考え方は現在の生態学の観点からは理解しやすいが、具体的な証拠としての化石のないのが難点である。
|
2─4 現在の動物界
現在の動物界は後生動物の約30の門からできている。研究者によって門の数に多少の違いがあるが、それはいくつかの分類群を独自の門とするか近縁の門の綱とするかの見解の違いによるもので、動物界には他とはっきり区別できる約30の大きな分類群があるという分類学上の見解は共通のものと言ってよい。しかし、これらの門の間の系統関係については、表現型にもとづく従来の見解と遺伝子型にもとづく近年の見解との間にまだ大きな相違がある。
表現型にもとづく系統関係は、その根拠に不確かな点の残っているものもあるが、100年を超える長い期間にわたって、発生学、形態学、生理学、生態学など多くの分野で蓄積された膨大なデータにもとづき、多くの研究者が議論を重ねて考察してきた。現在も広く用いられている表現型上の系統関係には、たとえば発生初期の原腸胚(嚢胚
gastrula)の原口(blastopore)が後の幼生および成体の口になる動物門を前口動物(旧口動物 Protostomia)とし、原口が後の幼生および成体の肛門になる動物門を後口動物(新口動物
Deuterostomia)としてまとめた例がある。また前口動物の中で、体表の上皮組織(epithelium)である外胚葉(ectoderm)と消化管の上皮組織である内胚葉(endoderm,
entoderm)の間に中胚葉(mesoderm)の上皮に囲まれた腔所(真体腔 deuterocoel)ができる真体腔動物(Deuterocoelomata,
Eucoelomata)と、中胚葉に囲まれない腔所(擬体腔 pseudocoel:胞胚腔由来)ができる擬体腔動物(Pseudocoelomata:袋形動物門
Phylum Aschelminthes として一括されたことがある)、およびどちらの腔所もできない無体腔動物(Acoelomata)をそれぞれまとめた例がある。そして後口動物は、みな真体腔をもつ真体腔動物であり、前口動物の真体腔動物から分かれたものと考えられた。これらは主に発生過程や形態上の共通点にもとづいて門の間の系統関係を推定した例である。
一方遺伝子型にもとづく系統関係は研究の歴史が浅く、系統解析に用いられる遺伝子 DNA あるいはその転写産物である RNA の種類は限られており、それらのデータの得られている生物種の数はまだ少ない。しかし、少ないデータから一定の信頼性を得るためのいくつかのプログラムが開発され、それらによって表現型からの結論とは必ずしも一致しない重要な系統上の見解が提示されている。たとえば脱皮動物説(Ecdysozoa
theory)では、従来の前口動物のうち表現型からは擬体腔動物に含まれる線虫(線形)動物門、線形虫(類線形、ハリガネムシ)動物門、動吻動物門、エラヒキムシ動物門、被甲(胴甲)動物門が、真体腔動物に含まれる節足動物門とともに単系統群になり、脱皮(ecdysis,
moulting)して成長するという共通の表現型(表現型による系統解析では硬いクチクラをもったことによる収斂形質とみなされている)にもとづき、脱皮動物(Ecdysozoa)と名づけられた。また脱皮動物以外の前口動物も同じく真体腔の有無にかかわらず単系統群になり、必ずしも全部には共通しない表現型の特徴(成体が触手冠
lophophore をもち、幼生がトロコフォア(trochophore、担輪子)型であるという特徴)にもとづき、冠輪動物(Lophotrochozoa)と名づけられた。
本講では、表現型上の多くの特徴にもとづいて現在の動物界の全体像を提示する。そこでは遺伝子型のデータを用いれば異なる結果が得られるような系統関係も、あるいは分岐論によれば側系統群になるかもしれない分類群も、表現型上の根拠があればとりあえず認めておく。それは1つには、本講の動物系統分類体系は、遺伝子型のデータと分岐論による新しい系統分類体系との比較のためにあえてここに提示するものだからであり(「まえがき」参照)、また今1つには、表現型にもとづく系統関係と遺伝子型にもとづく系統関係との間に不一致の生じる理由が、表現型からはもちろん、遺伝子型からもまだよく説明されていないと考えられるので、系統関係の構築における表現型上の多くの根拠をここで捨ててしまうのは少し早いのではないかと思うからである。しかし現在の学生諸君の便利のために、新しい見方を必要に応じてとり入れ、あるいは注記しておく。
ーー次回に続くーー
Contributed by Toshiki Makioka, Received September 19, 2006. Revised version received November 30, 2006.
©2006 筑波大学生物学類
|