![]()
![]()
![]()
![]()
![]()

渦鞭毛藻類は海洋から湖沼まで広い範囲に生息する鞭毛藻類で,その独特の形態で容易に他と区別できる。名前のdinoはギリシャ語のdines (=whirling, turning, 渦巻き,回転の意味)からきたもので,回転しながら泳ぐ姿に由来しています。
![]() 渦鞭毛藻の細胞はテカ(theca)またはアンフィエスマ(amphiesma)とよばれる独特の細胞外被構造をもっています。植物では一般に細胞膜の外に細胞壁などの外被構造があり,細胞を被っています。しかし,一部の藻類では細胞の形態を保持する外被構造が細胞膜の内側に発達する場合があります。渦鞭毛藻のアンフィエスマはその例で,細胞膜の内側に小胞があり,そのなかにセルロースからなる板状の構造(amphiesmal
plates, thecal plates)(「よろい」に似ていることから鎧板ともよばれる)が形成されます。このプレートの形と数,発達の程度および配列は種によって一定で,渦鞭毛藻の独特な外観が分類上の重要な性質とされています。
渦鞭毛藻の細胞はテカ(theca)またはアンフィエスマ(amphiesma)とよばれる独特の細胞外被構造をもっています。植物では一般に細胞膜の外に細胞壁などの外被構造があり,細胞を被っています。しかし,一部の藻類では細胞の形態を保持する外被構造が細胞膜の内側に発達する場合があります。渦鞭毛藻のアンフィエスマはその例で,細胞膜の内側に小胞があり,そのなかにセルロースからなる板状の構造(amphiesmal
plates, thecal plates)(「よろい」に似ていることから鎧板ともよばれる)が形成されます。このプレートの形と数,発達の程度および配列は種によって一定で,渦鞭毛藻の独特な外観が分類上の重要な性質とされています。
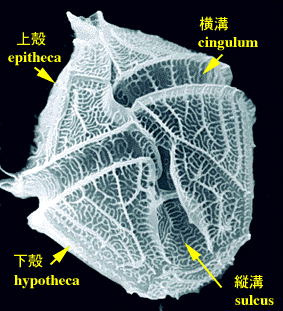 渦鞭毛藻の多くの種では,テカは上殻(epitheca)と下殻(hypotheca)とよばれる二つの部分からなります。上殻と下殻の間に横溝(cingulum)があり,また横溝と交差して細胞後方に縦溝(sulcus)が伸びています。典型的な渦鞭毛藻では上殻と下殻の大きさはほぼ同じ大きさですが,種によって上殻が極度に退化した(または未発達な)もの(Prorocentrum,
Ornithocercus, Amphisoleniaなど),下殻が退化した(または未発達な)もの(Katodiniumなど)があります。
渦鞭毛藻の多くの種では,テカは上殻(epitheca)と下殻(hypotheca)とよばれる二つの部分からなります。上殻と下殻の間に横溝(cingulum)があり,また横溝と交差して細胞後方に縦溝(sulcus)が伸びています。典型的な渦鞭毛藻では上殻と下殻の大きさはほぼ同じ大きさですが,種によって上殻が極度に退化した(または未発達な)もの(Prorocentrum,
Ornithocercus, Amphisoleniaなど),下殻が退化した(または未発達な)もの(Katodiniumなど)があります。
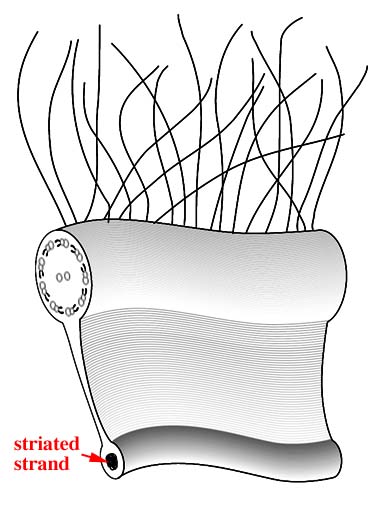 横溝と縦溝の交点に小型のプレートからなる部分があり,鞭毛孔を取り囲んでいます。そこから2本の鞭毛が生じます。1本は横鞭毛(transverse
flagellum)とよばれ,横溝に沿って伸びて細胞を取り囲み,他の1本は縦鞭毛(longitudinal
flagellum)で縦溝に沿って細胞の後方に伸びています。横鞭毛はらせん状にねじれて見えますが,これは通常の9+2構造をもつ鞭毛の本体(軸糸)とは別に,鞭毛を懸垂する繊維があるためです。この繊維はカルシウムイオンの濃度が上がると収縮するタンパク質(セントリン)を含んでおり,収縮の結果,運動の停止や遊泳の方向が変わります。横鞭毛には細い毛状の構造がみられますが,黄色植物の前鞭毛に普遍的にみられる管状マスチゴネマのように中空ではなく,また推進力の方向を逆転することもありません。
このような形態の鞭毛は他に例がなく,渦鞭毛藻を特徴づける重要な性質のひとつとなっています。
横溝と縦溝の交点に小型のプレートからなる部分があり,鞭毛孔を取り囲んでいます。そこから2本の鞭毛が生じます。1本は横鞭毛(transverse
flagellum)とよばれ,横溝に沿って伸びて細胞を取り囲み,他の1本は縦鞭毛(longitudinal
flagellum)で縦溝に沿って細胞の後方に伸びています。横鞭毛はらせん状にねじれて見えますが,これは通常の9+2構造をもつ鞭毛の本体(軸糸)とは別に,鞭毛を懸垂する繊維があるためです。この繊維はカルシウムイオンの濃度が上がると収縮するタンパク質(セントリン)を含んでおり,収縮の結果,運動の停止や遊泳の方向が変わります。横鞭毛には細い毛状の構造がみられますが,黄色植物の前鞭毛に普遍的にみられる管状マスチゴネマのように中空ではなく,また推進力の方向を逆転することもありません。
このような形態の鞭毛は他に例がなく,渦鞭毛藻を特徴づける重要な性質のひとつとなっています。
縦鞭毛(longitudinal flagellum)
縦鞭毛は
渦鞭毛核(dinokaryon, dinokaryotic nucleus)
そのほかにも渦鞭毛藻類の細胞は多くの特異な特徴をもっています。中でも最も注目するべきものは染色体です。
![]()
![]()
渦鞭毛藻の核では,静止期の細胞でも染色体がみられ,
生活環を通して染色体が凝集して存在しています。
真核生物のほとんどでは,DNAはヒストンと結合している。しかし,・・・・
真核生物のほとんどでは核DNAは塩基性タンパク質のヒストンと結合しています。ヒストン
(H2A, H2B, H3, G4)は8量体を形成し,それにDNAの二重鎖が巻き付いてヌクレオソーム
(nucleosome)がつくられます。ヌクレオソームはヒストン1によって結びつけられて染色質(クロマチン)をつくり,それがさらに巻きとられて染色体が作られている。渦鞭毛藻ではヒストン(なかでもヌクレオソームの架橋構造をつかさどるH1)はなく,代わりに類似の塩基性タンパクが結合しています。このように渦鞭毛藻類の核は真核生物の中でも大変ユニークで,かつては原核と真核の中間であるともいわれ,中核(mesokaryotic)などともよばれていました。しかし,渦鞭毛藻は進化的にはそれほど古い生物ではないと考えられます(化石は中生代以降から産する)。18SrDNのA分子系統樹でも渦鞭毛藻類がいわゆるクラウン生物群の一つであることが示唆されています。このことは,ヒストンを欠く渦鞭毛藻の核は二次的に生じたものであることを示しています。
二次的な消失とはいえ,真核生物のなかでヒストンを欠くことは極めて珍しいことで,渦鞭毛藻を特徴づけ重要な性質と言えます。
渦鞭毛藻の核の分裂は大変変わったものです。一般に知られている核の分裂は,中心体が複製されて分裂の二つの極に移動する,染色質が凝集して染色体が出現し,核膜が消失する。染色体は中心体の周辺から生じる微小管によって連結され,これが紡錘体として働いて両極に分離する,というものです。
しかし渦鞭毛藻では核の分裂の全期間にわたって核膜はいっさい消失することがありません。分裂に先立って中心体が複製され,両極に分かれます。この点では多くの真核生物と同様です。しかし,核膜は消失せず,両極を結ぶ微小管はやがて核に沈み込んでいって,核膜によって包み込まれます。結果として核を貫く細胞質のトンネルの中に微小管が走っているといった構造になります。染色体は核膜を介して紡錘体として働く微小管につながっており,やがて微小管の運動によって染色体は核膜に沿って両極に分かれていきます。
 渦鞭毛藻のおよそ半数は食作用などの従属栄養によって生活しています。残りの半数が光合成を行います。
渦鞭毛藻のおよそ半数は食作用などの従属栄養によって生活しています。残りの半数が光合成を行います。
渦鞭毛藻の要領体はさまざまなものが知られています。これは,多くの藻類の仲間で葉緑体のもつ性質が安定であることを考えると,不思議なことです。
渦鞭毛藻でもっとも普通にみられる葉緑体は次のような性質をもっています。
色素はクロロフィルaとc2および渦鞭毛藻特有のカロチノイドであるペリディニン(peridinin)で,これらが複合体を形成して光受容の役割を果たしています。ペリディニンをもつために,葉緑体の色彩は黄色植物(主要なカロチノイドはfucoxanthin)に比べると茶色がかっています。
葉緑体を包む膜は3枚です。3枚の膜をもつ葉緑体は渦鞭毛藻とユーグレナ植物だけで知られており,細胞共生と関連づけて説明されることが多いのですが,膜の起源については推測の域を出ず,不明な点が残されています。ラメラは3重チラコイドです
![]() 藻類の葉緑体(主要色素と形態)
藻類の葉緑体(主要色素と形態)
渦鞭毛藻のなかには,光合成色素として黄色植物とおなじフコキサンチンをもっている仲間があります。
驚くべき事に,クロロフィルaとbをもっている渦鞭毛藻もいくつか知られています。
![]() 目以下の渦鞭毛藻の分類で最も重要な性質が鎧板の配列のパターン
(thecal pattern)で,一定の配列をしていることから,しばしばtabulationとよばれます。鎧版が明瞭なペリディニウム目やゴニオラックス目の藻類では,この鎧版の配列パターンによって分類が行われています。
目以下の渦鞭毛藻の分類で最も重要な性質が鎧板の配列のパターン
(thecal pattern)で,一定の配列をしていることから,しばしばtabulationとよばれます。鎧版が明瞭なペリディニウム目やゴニオラックス目の藻類では,この鎧版の配列パターンによって分類が行われています。
Prorocentroid
テカは二枚貝の殻のような2枚の大型のプレートからなり,両者はかみ合って密着しています。鞭毛の出る細胞先端部では8-12個の小型のプレートが一定のパターンで配置されています。
Dinophysoid
Gymnodinoid
Gonyaulakoid
Peridinioid
渦鞭毛藻では,現在でも真核共生のさまざまな例,さまざまな段階をみることができます。光合成を行う大部分の渦鞭毛藻はペリディニン(peridinin)というキサントフィルをもっています。これは渦鞭毛藻に特有の光合成補助色素です。ところが,一部の渦鞭毛藻はペリディニンを欠き,代わりに黄色植物と同じフコキサンチンをもっています。Peridinium
balticumがその例です。また19'ヘキサノイルオキシフコキサンチンというハプト藻と同じ光合成色素をもつものもあります。これらは珪藻やハプト藻が渦鞭毛藻に取り込まれて葉緑体になったものと考えられます。
さらに,Gymnodinium acidotumやAmphidinium latum, Amphidinium
wigrenseなどの渦鞭毛藻はクリプト藻と同定される共生体をもっています。前二者ではクリプト藻の核,ヌクレオモルフが残っており,このことからクリプト藻を食作用によって外界から取り込み,消化することなく葉緑体として利用していると考えられています。それに対して,A.
wigrenseではクリプト藻の核もヌクレオモルフも退化しており,取り込まれた細胞が葉緑体として定着しているようです。
| Gymnodinium acidotum クリプト藻Chroomonasを共生体にもつ | |
| Amphidinium latum クリプト藻を共生体にもつ image courtesy of Takeo Horiguchi, Hokkaido Univ. 写真提供:堀口健雄(北海道大学) |
単細胞の生物には細胞から外界に向かって放出されるさまざまな射出構造が知られています。渦鞭毛藻でもトリコシスト(trichocyst)とよばれる構造があり,独自の形態をもつことでしられています。
多様な共生の結果として,渦鞭毛藻はさまざまな葉緑体をもった種を生み出しているが,渦鞭毛藻自体も他の生物に共生して「光合成を行う動物」を作り出している。このような「光合成動物」の中で最もよく知られ,地球環境問題の観点からも注目されているのがサンゴである。サンゴにはSymbiodinium microadriaticum (Zooxanthella microadriatica)とよばれる渦鞭毛藻が共生(この場合は細胞内ではなく,細胞間隙に生息)している。Symbiodiniumは,サンゴの他,貝やイソギンチャク,有孔虫など多様な生物と共生関係にある。
渦鞭毛藻はさまざまな独自の性質を持ち,他の生物の仲間との類縁がよくわからなかった藻類のひとつです。しかし,細胞微細構造と分子系統の研究から,「原生動物」の繊毛虫(ゾウリムシやラッパムシなど)や寄生性の微胞子虫類の仲間であるアピコンプレクサ(Apicomplexa)類に近縁であることがわかってきました。これらは細胞の表面構造が類似していること以外は生活の様式やすみかなど,互いに著しく異なる生物群です。このように一見大変かけ離れた生物が系統的にはひとつの仲間であることが明らかになることで,「藻類」のもつ多様さと生物界の認識における重要度が改めて認識されるようになってきました。