![]()
![]()
![]()
![]()
![]()

| 鞭毛は細胞の窪み(flagellar pitとよばれる)から生じる。 | |
| 鞭毛は多くの緑色藻類と異なり,先端は先細り(hair point)にならず,ひも状(truncate)である。 | |
| 鞭毛の表面にはゴルジ体で形成される鱗片が規則正しく配列されている。 | |
| ほとんどの種は細胞の表面も鱗片(scales)で被われる。 |
プラシノ藻は緑色植物の初期進化を探る上で極めて重要な生物群と考えられる。プラシノ藻の系統が明らかになれば,緑色植物の分類のかなりの部分が理解されると思われます。
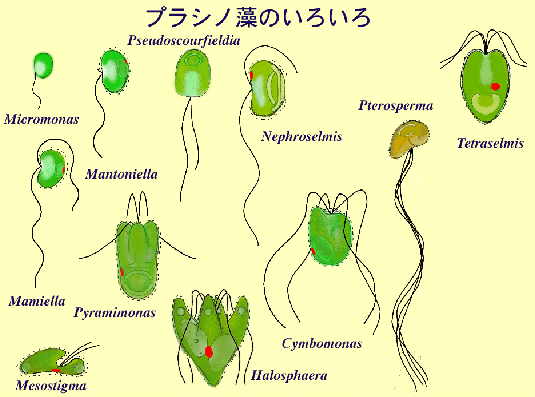
図 プラシノ藻のいろいろ
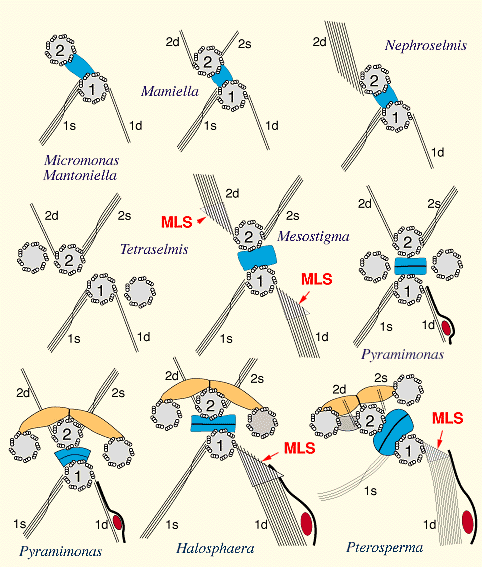 プラシノ藻の多様性は鞭毛装置にも表れている。他の緑色植物(緑藻,アオサ藻,シャジク藻)の鞭毛装置がそれぞれほぼ一様であることに対して,プラシノ藻では鞭毛装置も多様性に富んでいる。
プラシノ藻の多様性は鞭毛装置にも表れている。他の緑色植物(緑藻,アオサ藻,シャジク藻)の鞭毛装置がそれぞれほぼ一様であることに対して,プラシノ藻では鞭毛装置も多様性に富んでいる。
なかでも,Mesostigma, Halosphaera, Pterospermaなどの属では,交叉型の鞭毛根とともに,1dの鞭毛根に多層構造体(MLS)が付随しており,緑藻・アオサ藻群(交叉型鞭毛根で特徴づけられる)とシャジク藻・陸上植物群(多層構造体の存在で特徴づけられる)の双方の性質を合わせ持っている。
ほとんどのプラシノ藻の細胞と鞭毛は鱗片(scale)で被われている。鱗片はゴルジ体で形成され,鞭毛の基部から細胞外に放出される。鱗片の形態は属や種で一定で,プラシノ藻を同定する分類形質となっている。
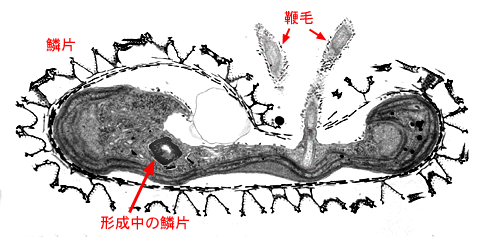 Mesostigma(細胞と鞭毛表面は鱗片に被われている)
Mesostigma(細胞と鞭毛表面は鱗片に被われている)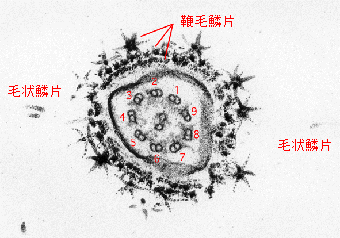 鞭毛鱗片(flagellar
scales)には鞭毛表面を層をなして密に被う鱗片と,両側に列をなす毛状鱗片(hair
scale)がある。最下層のダイアモンド鱗片は直径約40nmの4または5角形の鱗片で,マミエラ目以外のすべてのプラシノ藻に存在する。ダイアモンド鱗片のは分類群によって異なる形態(たとえばピラミモナス目ではカブトガニの形)の鱗片で被われている。Nephroselmisではさらに3層目,4層目の鱗片が知られている。
鞭毛鱗片(flagellar
scales)には鞭毛表面を層をなして密に被う鱗片と,両側に列をなす毛状鱗片(hair
scale)がある。最下層のダイアモンド鱗片は直径約40nmの4または5角形の鱗片で,マミエラ目以外のすべてのプラシノ藻に存在する。ダイアモンド鱗片のは分類群によって異なる形態(たとえばピラミモナス目ではカブトガニの形)の鱗片で被われている。Nephroselmisではさらに3層目,4層目の鱗片が知られている。
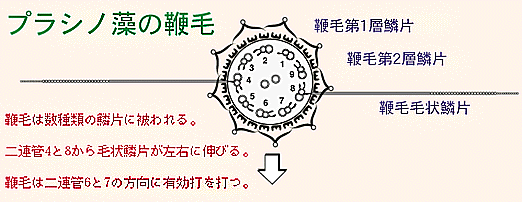 これらの鱗片は鞭毛上で特定のパターンで配置されている。毛状鱗片は鞭毛の有効打(effective
stroke)の向きに対して直角の方向に伸びており,鞭毛の推進力の増加に寄与していると考えられる。9つの2連管のうちの一つはダイニン外腕を欠いている。この2連管を1番と名づけ,鞭毛先端からみて反時計回りに番号をふることができる。このとき,有効打の方向は6番の方向にあたり,また毛状鱗片は4番と8番の2連管から左右に伸びている。
これらの鱗片は鞭毛上で特定のパターンで配置されている。毛状鱗片は鞭毛の有効打(effective
stroke)の向きに対して直角の方向に伸びており,鞭毛の推進力の増加に寄与していると考えられる。9つの2連管のうちの一つはダイニン外腕を欠いている。この2連管を1番と名づけ,鞭毛先端からみて反時計回りに番号をふることができる。このとき,有効打の方向は6番の方向にあたり,また毛状鱗片は4番と8番の2連管から左右に伸びている。
Melkonian (1989の分類系)
| Mamiellales (マミエラ目) |
Mamiella, Dolicomastix, Mantoniella, Bathycoccus, Micromonas |
| Pseudoscourfieldiales (シュードスコールフィールディア目) |
Pseudoscourfieldia, Nephroselmis, Picnococcus |
| Chlorodendrales (クロロデンドロン目) |
Tetraselmis, Scherffelia |
| Pyramimonadales (ピラミモナス目) |
Pyramimonas, Cymbomonas, Pterosperma, Pachysphaera, Halosphaera, Mesostigma* |
{kind=link}
{kind=link}