1,細胞壁マトリックス糖鎖合成に必須である糖ヌクレオチド関連遺伝子の胚発生・花形成における機能
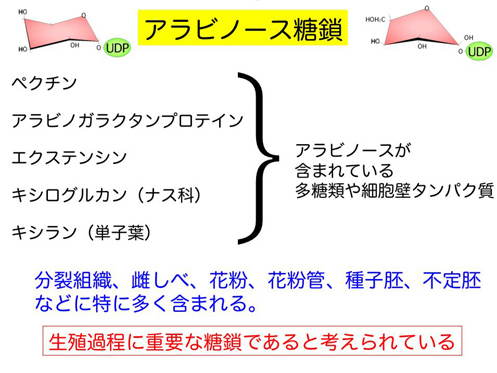
高等植物の生殖過程には、細胞壁マトリックス糖鎖を介した細胞接着が様々な場面で要求される。しかし、そのメカニズムと機能に関する知見は極めて乏しいのが現状である。現在までに、半数体タバコ培養系による細胞接着変異体作出系を開発することで、ペクチン側鎖合成に関わるアラビノース鎖合成関連遺伝子が、現在までに同定している。また、それと協調して働くと考えられる新規アラビノースムターゼ遺伝子UAM(UDP-Arabinose mutase)が、共同研究者である森林総合研究所の石井忠博士により同定された。アラビノースムターゼは、熱力学的には安定なアラビノピラノース(六員環)を実際の細胞壁糖鎖として利用されているアラビノフラノース(五員環)へと転換する活性を有している酵素である。アラビノース糖鎖は、高等植物が特徴的に有する糖鎖でありアラビノガラクタン蛋白質をはじめ、生殖過程に大きな関わりを持つ糖鎖である。また、これらは生殖過程における強い発現が確認されている。これらのペクチン糖鎖関連遺伝子は高等植物間で90%以上相同性を持つ保存性の高い遺伝子であり、その機能も広く保存されていることが期待できる。また、アラビドプシスUAM1, 2の二重変異体では、不稔になることが確認され、生殖器官の形成などにも重要であることが示唆されている。本研究では、いくつかのアラビノース合成関連遺伝子の解析を解明することで、アラビノースを有する細胞壁マトリックス糖鎖が花粉形成および花粉管ガイダンス、そして胚発生・種子休眠&発芽において果たす役割について分子レベルで解明することを目的としている。
 |
| 琉球大学
小西照子先生 |
2,双子葉植物および単子葉植物における細胞壁マトリックス関連遺伝子の機能
1)タバコを用いた研究
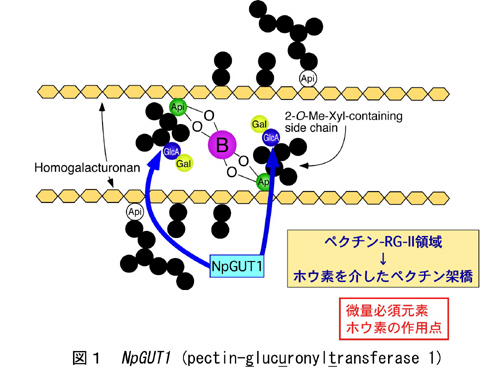
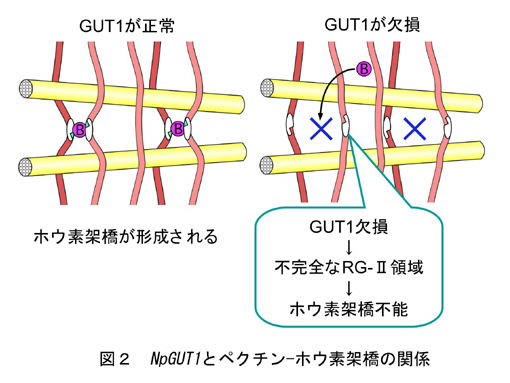
多細胞生物である高等植物の形態形成において、細胞自身の分化に加え、分化した細胞同士の接着が重要である。しかし、その細胞接着の主役であるペクチン関して,その生合成メカニズムや発現特性に関する知見は極めて乏しい。近年我々は、Nicotiana plumbaginifolia の半数体植物の葉切片にT-DNAを挿入して培養し、不定芽形成能力を失うと同時に、細胞接着性の弱くなった突然変異体nolac (non-organogenic callus with loosely attached cells) の作出法を確立した(文献1)。この系を用いれば、シロイヌナズナでは作出が困難な胚性致死の細胞接着変異体をカルスとして維持し、変異体の生化学的解析を行うことが可能となる。そのうちの一つであるnolac-H18の解析より、新規ペクチングルクロン酸転移酵素遺伝子(NpGUT1; glucuronyltransferase 1)が同定された(文献2)。この遺伝子は、植物のペクチン合成に関わる初めての糖転移酵素遺伝子で(図1)、頂端分裂組織で特に発現が強く、メリステム形成と共にホウ素の作用点であるラムノガラクツロナンII(RG-II)二量体の形成に必須であることが判明している(図2)。
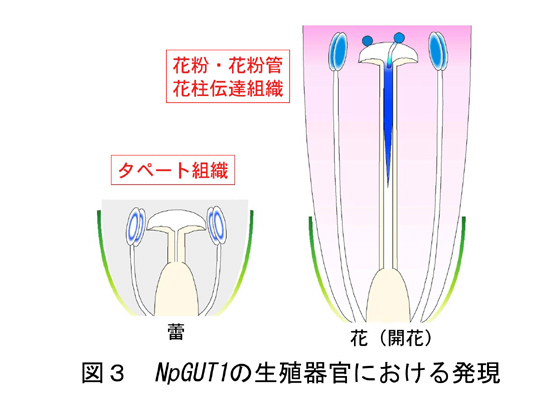
NpGUT1は、栄養器官においては茎頂及び根端の分裂組織において発現するが、生殖器官における詳細な発現を、pNpGUT1::GUS形質転換タバコおよびin situ hybridization法を用いて調査したところ、タペート組織、花粉、花粉管の先端、花柱の伝達組織において発現が見られた(図3)。以上の結果よりNpGUT1は、分裂組織における細胞接着に重要であるだけでなく、花粉管ガイダンスにも重要である可能性が示唆された。これらの発現をDEX誘導性プロモータ-アンチセンスNpGUT1を導入することにより抑制し、受精に関わる組織の形成や受精プロセスが阻害されるか検討した。
未熟な雄蕊においてDEXによりNpGUT1の発現を抑制したところ、成熟した葯には正常な花粉が形成されず、葯の中はほとんど空洞であった。花粉をDEX存在下において発芽させたところ全く発芽しなかった。また発芽花粉にDEXを与えると、花粉管伸長が著しく阻害された。NpGUT1の発現抑制によって伸長抑制の起きた花粉管に対して、抗RG-II抗体を用いてのRG-IIの局在および、レーザーアブレーションICP-MSによるホウ素の定量分析を行った。その結果、発現抑制による伸長抑制が起きた花粉管の先端では、RG-II、ホウ素ともに消失していることが確認された(図4)。
一方、未熟な雌蕊にDEXを処理したところ、成熟したほぼ全ての花柱において伝達組織に異常が生じ、不稔となった(図4)。NpGUT1の発現抑制による花柱の伝達組織の形成阻害が起きた雌しべが、花粉管を誘導できるかどうかをアッセイしてみたところ、花粉管は花柱の中を通過できなかった(文献3)。
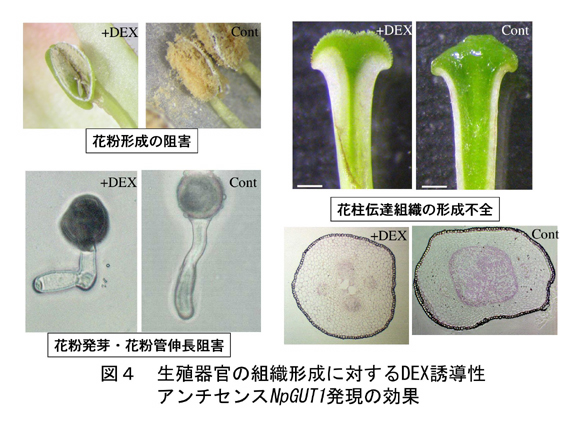
NpGUT1は、ペクチンのホウ素を介した架橋の形成に重要であることが明らかとなっているが、この遺伝子はメリステム等における細胞接着のみに機能するのではないことが明らかとなった。花粉の形成、花粉管の伸長、さらには花柱伝達組織の形成といった、細胞分化や細胞伸長にも関わっている可能性が示唆された。これは、ペクチンが必須微量元素であるホウ素の主要な受容体であることと関連していると考えられる。また、花粉管と花柱伝達組織との間における接着やガイダンスにも関わっている可能性が高い。これらの事実から、NpGUT1の発現調節により、雄性不稔誘導や受精の調節ができる可能性が期待される。
2)イネを用いた研究
現在、細胞壁マトリックスの合成および分解を調節したイネを作成し、花器官の発生および受粉、花粉管ガイダンス、受精について調査を行っている。
 |
住吉 美奈子
|
生命環境科学研究科
生命共存科学専攻3年 |
単子葉植物におけるアラビノースネットワーク構築機能の解明
|
 |
四谷 紗和子
|
生命環境科学研究科
生命共存科学専攻2年 |
単子葉植物におけるペクチン-ホウ素架橋関連遺伝子の機能解明
|
 |
市川 愛
|
生命環境科学研究科
生命共存科学専攻2年 |
単子葉植物の生殖過程における細胞壁マトリックス糖鎖の機能解明
|
 |
武部 尚美
|
生命環境科学研究科
生命共存科学専攻2年 |
単子葉植物におけるグリシンリッチタンパク質の機能解明
|
 |
稲村 拓也
|
生命環境科学研究科 生命共存科学専攻2年 |
単子葉植物におけるアラビノース合成メカニズムの解明
|
 |
宮下 彩 | 生物学類4年 | 単子葉植物におけるプロリンリッチタンパク質の機能解明 |
 |
長谷川 和也 | 生物学類4年 | イネにおけるペクチン分解関連遺伝子阻害タンパク質の機能解明 |
 |
小原 崇司 | 生物学類4年 | 植物の病害応答における細胞壁の機能について |
参考文献
1) H. Iwai, T. Ishii and S. Satoh (2001) Absence of arabinan in the side chains of the pectic polysaccharides strongly associated with cell walls of Nicotiana plumbaginifolia non-organogenic with loosely attached constituent cells. Planta 213: 907-915.
2) H. Iwai, N. Masaoka, T. Ishii and S. Satoh (2002) A pectin glucuronyltransferase gene is essential for intercellular attachment in the plant meristem. Proc Natl Acad Sci USA 99 (25): 16319-16324.
3) H. Iwai, A. Hokura, M. Oishi, H. Chida, T. Ishii, S. Sakai and S. Satoh (2006) The gene responsible for borate cross-linking of pectin RG-II is required for plant reproductive tissue development and fertilization. Proc Natl Acad Sci USA 103 (44): 16592-16597.
4) H. Iwai, T. Ishii, S. Sakai and S. Satoh (2006) Molecular and Functional Analysis of Pectins in Intercellular Attachment by T-DNA Tagging Using a Haploid Tobacco Tissue Culture System. The Science and Lore of the Plant Cell Wall Biosynthesis, Structure and Function, Brown Walker Press 242-248.
・岩井宏暁、佐藤忍 (2002) 細胞接着とペクチン生合成 蛋白質・核酸・酵素 47 (12): 1618-1619
・新聞報道:2006年 10月23日 読売新聞