|
つくば生物ジャーナル Tsukuba Journal of Biology (2003) 2: 4-5.
特集:生物多様性
生物多様性研究の現在
中山 剛
(筑波大学 生物科学系)
我々の身のまわりにはさまざまな生物が生きており、直接・間接に人の生活に関わっている。私たちはこの生物 たちの生き様を学び、さまざまな切り口から研究を行っているが、その際には生物の多様性をできるだけ正しく理解することが必要である。生物間の相互の歴史的関係(系統)を知ることは、人の知的好奇心の大きな対象であると同時に、全ての生物学分野における基礎情報として不可欠である。ここでは最近の話題を含めて生物界全体の系 統的多様性について概説したい。
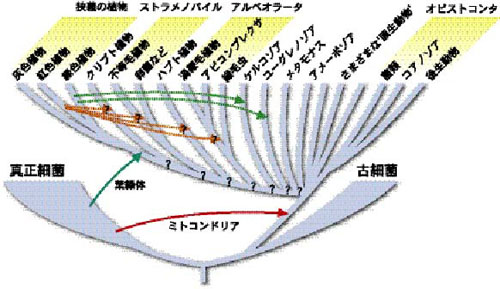
◆ 古細菌と真核生物古くは生物を大きく植物と動物に二分していたが、生物界における最大の不連続は原核生物と真核生物の間にあることはご存じの通りである。ただし系統的に見てみると、原核生物の中には2つの大きなグループ、真正細菌と古細菌があり、後者が真核生物に近縁であるらしい(図参照)。最近では環境中のDNAを直接採取して系統解析を行うことが一般的になり、いまでは原核生物のそれぞれのグループ内での多様性はこれまでの認識を遙かに上回るものであることがわかっている。特に古細菌では界レベルで既知のものとは異なる系統群が見つかっており、真核生物の起源を考える上でも古細菌の系統的多様性の解明は今後重要になってくるだろう。
生物全体の模式的な系統樹
実線の矢印は一次共生を、破線の矢印は二次共生を示す。真核生物内のそれぞれのグループ間の関係や葉緑体の二次共生の回数については不明な点も多い。
◆ 真核生物の多様性我々ヒトや陸上植物を含む真核生物という大きな生物群の多様性についてかなり理解が進んだ。この多様性の理 解を助けたのが「細胞内共生」という考えである。独自のDNAや70S型リボゾームの存在などから、葉緑体やミト コンドリアが共生した原核生物起源であることは有力視されていたが、さまざまな遺伝子・遺伝的システムの研究 からもこの考えは支持され、現在では定説となっている。ミトコンドリアがα-プロテオバクテリアに起源をもつ ことは明らかであるが、どの段階で真核生物に共生したかについては最近まで二転三転していた。いわゆる原生動 物の中に、ランブル鞭毛虫や微胞子虫などミトコンドリアを欠くとされていたものがいる。初期の分子系統学的研 究では、これらの生物が真核生物の中で最初に分岐した生物であることが示され、ミトコンドリアを獲得する前の 真核生物であると考えられていた。このことからこれらの生物はアーケゾアとよばれていたが、最近になっていく つかのアーケゾアはミトコンドリアをもつ生物よりも後に分岐したことが詳細な系統解析から示されるとともに、多 くのアーケゾアはミトコンドリア起源の遺伝子を(あるものでは退化したミトコンドリアさえも)もつことが明ら かになってきた。これらのことから、現在ではアーケゾアはミトコンドリアを二次的に失った生物であり、全ての 真核生物はミトコンドリアを獲得した共通祖先に由来すると考えられている。
◆ 真の原生動物アーケゾアは解体されたが、最近の分子系統学的研究によってアメーボゾアやケルコゾアなどさまざまな原生動物の新たなグループが認識されてきた。これらのグループは系統的には多細胞動物(後生動物)や菌類、緑色植物に匹敵するようなまとまりであり、古典的な意味での原生動物はさまざまな系統の真核生物の寄せ集めだといえる。 その中で「真の原生動物」、つまり後生動物に近縁な生物群がコアノゾアである。コアノゾアに含まれる襟鞭毛虫類は、海綿動物の襟細胞との類似性から古くから後生動物との近縁性が示唆されており、分子系統解析からも支持されている。近年では襟鞭毛虫の他にも胞子虫的な生物(e.g. Ichthyosphonus)やアメーバ的な生物(e.g. Nuclearia)もコアノゾアに属することがわかってきた。また後生動物やコアノゾアは菌類に近縁であることも明らかになっており、このまとまりは細胞後端から生じる1本の鞭毛という共通した特徴からオピストコンタ (Opisthokonta)と名付けられている。重要な生物群である後生動物や菌類の起源やそこで見つかった現象の進化を知るためには、単細胞段階にあるコアノゾアに関する研究が重要になってくるだろう。
◆ 光合成真核生物葉緑体が共生した藍藻(シアノバクテリア)に起源をもつことは明らかであるが、その共生が何回起こったのか? という点についてはいくつかの説がある。特に葉緑体の場合、真核生物と原核生物の共生だけでなく、真核生物同士の共生も考慮しなければならない。前者の共生は一次共生、後者は二次共生とよばれるが、その進化的な違いは葉緑体の微細構造に残されている。一次共生の葉緑体は2枚の膜で囲まれているが、二次共生由来と考えられる葉緑体はその外側をさらに膜で覆われており、本来の葉緑体膜の外側には核に類似した構造が存在することもある。この核様構造はヌクレオモルフとよばれ、共生した真核生物の核の名残であると考えられている。最近ではこのヌク レオモルフのゲノム解析も進み、二次共生機構の解明が期待されている。このような複雑な歴史をもつ葉緑体の進化は、植物だけではなく真核生物全体の系統と密接に関連している。葉緑体ゲノムの研究が進むにつれ、葉緑体は藍藻には見られない共通の特徴を多く持つことが明らかになり、分子系統学的にも全ての葉緑体が単一の起源をもつことが明らかになっている。つまり一次共生はただ1回の事象だったのだ。このことは宿主の方からも確かめられており、一次共生起源の葉緑体をもつ生物、灰色植物、紅色植物、緑色植物は単系統群であることが核・ミトコンドリア遺伝子の系統解析から示唆されている。では二次共生の方はどうだろうか?二次共生による葉緑体をもつ生物群は6つが知られているが、それらは互いに全く異質な細胞構造をもつ。さらに多くの場合、それぞれのグループに葉緑体をもたない姉妹群が知られており、これら6つの生物群における2次共生は独立に起こったとするのが一般的である。ストラメノパイル(コンブやミズカビなど)やアルベオラータ(渦鞭毛藻、マラリア原虫、繊毛虫など)、ユーグレノゾア(ミドリムシ、眠り病原虫など)のような生態的に全く異なる、古典的な意味での植物・動物・菌類にまたがるような生物同士のまとまりは、このような二次共生を考慮することで理解できる。またアピコンプレクサ(マラリア原虫など)が光合成能を欠く葉緑体をもっているという事実は、藻類と原生動物の間のつながりを示すとともに、葉緑体のもつ役割を考えさせる現象である。 多数回の二次共生という考えは、上記のような真核生物の関係を理解する際に都合がいいが、近年これに反する報告がいくつかなされている。不等毛植物(コンブなど)、ハプト植物、クリプト植物の葉緑体は二次共生に由来するが、これらが単一の紅色植物に起源をもつということが最近示された。また渦鞭毛藻を含むアルベオラータと不等毛植物を含むストラメノパイルが近縁であることも報告されている。もしこれらの生物での二次共生が単一の現象であったならば、ミズカビやオパリナ、繊毛虫などはもともと葉緑体をもっていたものが二次的にこれを失ったと考えざるを得ない。さらにオピストコンタを除く多くの真核生物が系統的にひとまとまりであり、これらはもともと葉緑体をもっていたことを示唆する報告もあり、真核生物全体の進化像は今ダイナミックに変化している。一方で特に原生動物とされていた生物に関しては基礎的な研究が立ち遅れており、その系統的多様性のごく一部しか明らかになっていない。今後大きなレベルでの系統解析と同時に、基礎的な多様性の解明が進められていくことに よって、初めて真核生物の進化の全体像が明らかになっていくだろう。
Contributed by Takeshi Nakayama, Received January 16 2003, Accepted January 22 2003.
©2003 筑波大学生物学類
|