小胞子の形成
小胞子の形成
最初に葯室の中で胞原細胞 (archesporial cell) からなる胞原組織 (ふつう1層の細胞層) が形成される。胞原細胞は並層分裂をして、側膜細胞 (側壁細胞) からなる側膜細胞層 (parietal cell layer) と、胞子形成細胞からなる胞子形成組織 (sporogenous tissue) に分化する。
側膜細胞層は、内被 (endothecium) 、中間層 (middle layer) 、タペート組織 (タペータム tapetum) に分化する。内被はふつう1層、ときに多層の細胞層からなる。内被細胞の細胞壁は縞状に肥厚しており、おそらく葯の裂開に寄与している。中間層はふつう短命であり、花粉形成の過程で崩壊してしまう。中間層の細胞はデンプンなどに富んでおり、おそらく花粉形成に用いられる栄養分を貯蔵している。
側膜細胞層の最内層にあるタペート組織は、形成中の小胞子・花粉粒に接しており、花粉形成に対する栄養・原料供給に重要な働きを果たしている。タペート組織は基本的に側膜細胞層から形成されるが、部分的に葯隔組織から形成されることもある。起源が異なるタペート細胞が異なる形態を示す場合、二型性タペート組織という (Alectra [ゴマノハグサ科] など)。またキンギョソウ (ゴマノハグサ科) やホウセンカ (ツリフネソウ科) では胞子形成組織からタペート組織ができることがある。
タペート組織には以下の2型があるが、その中間型も存在する。
- 分泌型 (secretory type、腺型 glandular type、側膜型)
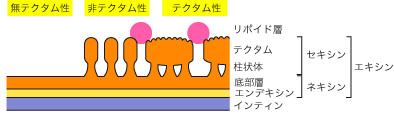
- タペート組織は本来の位置 (胞原組織の外側) にとどまり、葯腔 (葯室 locule) に物質を分泌する。タペート組織は次第に退化し、花粉成熟時には崩壊・消失する。分泌型タペート組織は、葯室にユービッシュ体 (Ubisch body) とよばれる構造を分泌するが、これはスポロポレニンからなり、花粉粒の細胞壁形成に関与しているらしい。イネ科、キンポウゲ科、ナデシコ科などに見られる。
- アメーバ型 (侵入型、周辺質型 amoeboid type)
- 比較的初期にタペート組織の細胞壁が消失し (自身の加水分解酵素による)、原形質が葯室にある花粉母細胞や花粉四分子の間に侵入する。タペート細胞は融合して多核の変形体 (plasmodium 周辺変形体 periplasmodium) となり、その中に花粉四分子が浮遊している状態になる。サトイモ科、ツユクサ科、メギ科、キク科などに見られる。
タペート組織の内側または外側には、タペート膜 (tapetal membrane) と呼ばれるアセトリシス耐性をもつ壁が形成される。タペート膜はタペート組織からの分泌物でできており、形成中の小胞子を取り囲んでいる。
タペート組織は花粉形成に大きく関与しており、花粉母細胞の減数分裂が始まる前に活性が高くなり、DNA量が著しく増加する。不完全なタペート組織は花粉の不捻性につながり、また花粉表面 (花粉外被) に存在するタペート組織起源のタンパク質は胞子体型自家不和合性に関与する (後述参照)。
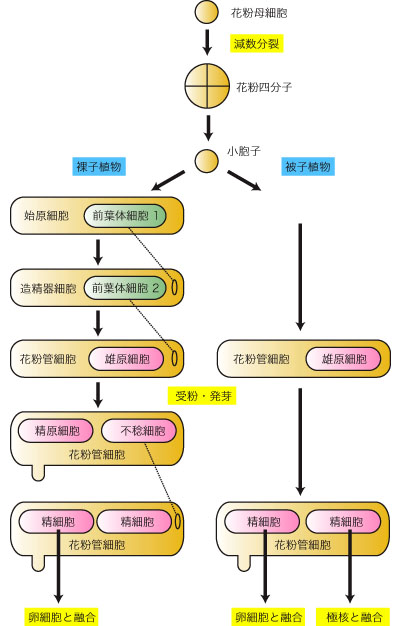
胞子形成組織の細胞は、直接または分裂を繰り返して花粉母細胞 (pollen mother cell, PMC 小胞子母細胞) となる。花粉母細胞はセルロース性の細胞壁をもち、互いにまたタペート細胞との間に原形質連絡をもっている。やがて花粉母細胞が減数分裂に入ると、細胞壁にカロースが沈着し、タペート細胞との原形質連絡は絶ち切られるが、花粉母細胞間の原形質連絡は太くなって (直径 1-2 µm) 細胞質チャネルになる。おそらく細胞質チャネルの存在によって、葯内の多数の花粉母細胞における減数分裂の同調性が保たれる。やがて減数第一分裂に入ると細胞質チャネルも断ち切られ、個々の花粉母細胞は独立した存在になる。ガガイモ科 (=キョウチクトウ科) やラン科のように花粉塊を形成するものでは、花粉母細胞細胞壁へのカロースの沈着が見られなかったり、細胞質チャネルが存続するなどの変異が見られる。
花粉母細胞は減数分裂を経て単相の小胞子 (microspore) が4個集まった花粉四分子 (pollen tetrad) を形成する。花粉母細胞の減数分裂には、大きく分けて以下の2型が見られる。
- 連続型 (successive)
- 減数第1分裂後に細胞質分裂、細胞壁形成 (カロース沈着を伴う) が起こって二分子が形成される。細胞壁形成は遠心的である。個々の二分子では減数第2分裂が起こるが、これは必ずしも同調しない。結果として単相の花粉四分子が形成される。
- 同時型
- 減数第1分裂後に細胞質分裂が起こらず、2核の細胞が形成される。2核は同調して減数第2分裂を行い、その後カロース沈着を伴った細胞壁が求心的に形成される。結果として単相の花粉四分子が形成される。
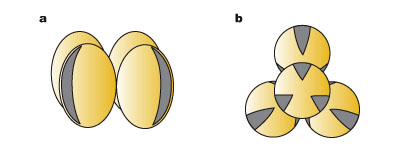
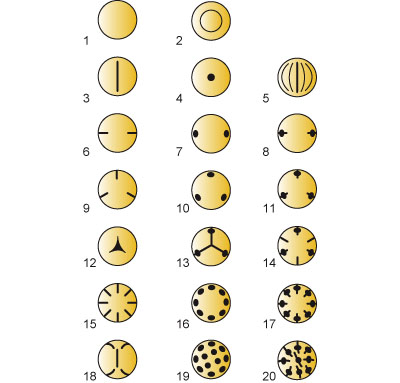
上記のような花粉四分子の形成過程の多様性は、結果としてできた花粉四分子における小胞子の配置にも影響し、また形成される小胞子 (→花粉粒) のタイプにも関係している (図2)。これら違いには系統的な意味があり、双同側型が原始的であって原始被子植物や単子葉類に見られ、四面体型が派生的で真正双子葉類の特徴になっている。
- 双同側型 (isobilateral)
- 基本的に連続型の花粉四分子形成によってでき、一平面上に4つの小胞子が列んでいる。形成される花粉は単溝粒を基本とする。
- 四面体型 (tetrahedral)
- 基本的に同時型の花粉四分子形成によってでき、四面体の各頂点に小胞子が配置されている。形成される花粉は三溝粒を基本とする。
花粉四分子における小胞子の配置には、少数ではあるが他にもいくつかのタイプが見られ、十字対生型、線状型、T字型などが知られている。
最終的に、タペート組織から分泌されたカレースによって花粉四分子のカロース壁は分解され、個々の小胞子が遊離した状態になるが、ツツジ科のように花粉四分子が集合したまま成熟・放出されるものもある (4集粒)。