 細胞壁
細胞壁
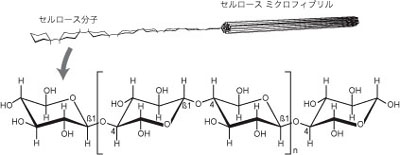
陸上植物の細胞は、せルロース (cellulose、β-1,4-D-グルカン) を主成分とした細胞壁 (cell wall) で囲まれている。細胞壁の存在は、陸上植物と多細胞動物の間の大きな違いの1つである。セルロースは直鎖状の高分子であり (図1)、同一方向に列んだ多数の分子が水素結合を介して束になることでセルロース微小繊維 (microfibril) を形成している。セルロース微小繊維はふつう互いに平行に列んで細胞壁の骨組みとなっている。セルロース量は植物体乾燥重量の1/3~1/2に達し、地球上の有機炭素の約半分はセルロースであるといわれる (年間生産量約1,000億トン)。セルロースは綿や紙などの形で人間にも身近な存在である。人間はセルロース分解酵素を持たないため、直接セルロースを栄養源とすることはできない。セルロース合成酵素は細胞膜上でロゼット型の複合体 (terminal comples; TC) を形成しており、細胞質側から供給される UDP-グルコースを原料にしてセルロースを合成し、細胞外に分泌する。
 セルロースについて
セルロースについて
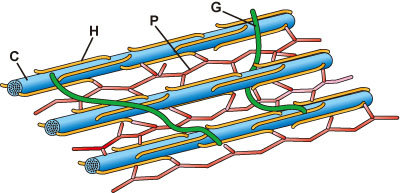
セルロース微小繊維の骨格は、ヘミセルロース (hemicellulose) (表1)、ペクチン (pectin) (表2)、タンパク質などからなるマトリックス (基質 matrix、基質ゲル matrix gel) の中に埋め込まれている (図2)。最近では、ヘミセルロースやペクチンなど非セルロース多糖を、マトリックス多糖 (matrix polysaccharide) と総称することも多い。タンパク質としてはエクステンシンのように細胞壁維持に働くものや、細胞壁多糖を分解して細胞の拡大成長に働くもの、細胞間認識に働くものなどが存在する。
ヘミセルロースについてペクチンについて細胞壁糖タンパク質について細胞壁酵素について
その他にも、細胞の種類によって、細胞壁にはさまざまな物質が蓄積している。導管細胞や繊維などの二次壁にはフェノール化合物であるリグニン (lignin) が多く蓄積し、細胞壁の機械的強度を増している。このような細胞壁を木化 (リグニン化 lignification) したという。リグニンはセルロースに次いで地球上に多い有機炭素化合物であるといわれる。また維管束などを取り囲む内皮細胞同士の接点 (カスパリー線) やコルク細胞には不飽和脂肪酸であるスベリン (suberin) が蓄積し、水や病原菌の侵入を防いでいる。さらに表皮細胞において外界に接する細胞壁には、不飽和脂肪酸であるクチン (cutin) や脂肪酸エステルのワックス (蝋 wax) が蓄積し、クチクラ層 (cuticular layer) を形成している (クチクラ化 cuticularization)。クチクラ層もまた、水の蒸発や病原菌の侵入などを防いでいる。表皮以外でも、植物体内において空気と接する細胞壁には薄いクチクラ層 (内部クチクラ interal cuticle) が存在する。ほかにも篩管細胞の連結部 (篩孔) が形成される際には β-1,3-グルカンであるカロース (callose) が重要な働きを演じ、篩管細胞が損傷を受けた際にもカロースによって篩孔がふさがれて栄養分の損失を防ぐ。また病原菌による壊死細胞周辺の細胞の細胞壁にもカロースが蓄積され、病原菌の感染拡大を妨げる。
陸上植物の細胞壁には、一次壁と二次壁があり、細胞壁同士は最外層の中葉で接着している。
- 一次壁 (primary wall)
- 一次壁は細胞が分裂してすぐにできる細胞壁である。一次壁はマトリックスの割合が多いため、伸縮性に富み、細胞の拡大成長・伸長成長にともなって細胞壁をある程度引き延ばすことができる。組成比は一般的にセルロース25~30%、ヘミセルロース25%、ペクチン30%、その他20%ほどである。また一次壁は水含量が大きい (重量比で~60%)。ミクロフィブリルの並びが内側では細胞長軸に直角であったものが外側に行くにつれ平行になるタイプ (マルチネット構造) と、直角の層と平行な層が互い違いに配列するタイプ (交差多層構造) がある。マルチネット構造は茎や根の皮層細胞で、交差多層型は茎の表皮細胞などに見られる。ふつう厚さ0.1~1 µmほどだが、厚角細胞や篩管細胞、表皮細胞などでは一次壁がかなり厚くなることが多い。
- 二次壁 (secondary wall)
- 拡大成長が終わった細胞において、一次壁の内側に形成される細胞壁。特に管状要素 (導管細胞や仮導管) や厚壁細胞などで顕著である。一次壁にくらべて強固で可塑性が低い。セルロースが一次壁より多く (約60%) 、ミクロフィブリルが数層 (ふつう3層) に列び、異方性が強い。またリグニンの蓄積によって機械的強度をさらに増している。二次壁は一次壁とは異なり、ペクチンやタンパク質含量が少なく、ヘミセルロースではキシランのような直鎖状分子の割合が多い。
- 中葉 (middle lamella)
- 隣接する細胞壁同士が接するところ、つまり一次壁の外側に存在し、ペクチンを主成分とする。中層または細胞間層 (intercellular layer) ともいう。細胞間接着などに関わっている。
細胞壁は外界からの異物の侵入を防ぐと共に、細胞を一定の形に保つ働きを果たしている。水など浸透圧の低い液体 (低張液) にさらされることによって細胞は膨らむ力、つまり膨圧 (turgor pressure) が生じ、これを細胞壁で押さえつけることによって組織の機械的支持力が得られる (だから水分が足りなくなると萎れてしまう)。また膨圧がある状態で細胞壁 (の一部) が細胞壁タンパク質の働きでゆるむことによって細胞の拡大成長が可能になる。
上記のようにリグニンやスベリン、クチン、蝋などが蓄積しない限り、細胞壁は水や低分子の物質を自由に通すことができる。このような細胞壁を含めた細胞外の経路を通じた物質輸送は、アポプラスト輸送 (apoplastic transport) とよばれ、原形質連絡を通じた原形質間の輸送であるシンプラスト輸送 (synplastic transport) と同様に植物体の物質輸送に重要な働きを果たしている。
細胞壁は静的な存在であり、非生物的な構造だと考えられがちである。しかし実際には細胞壁では活発な代謝が行われており、植物の本質的な生理機能に不可欠な存在である。ただし植物細胞の生存自体に細胞壁は細胞壁は必要不可欠ではない。花粉内の精細胞のように細胞壁を欠く細胞も存在するし、人為的に細胞壁を除去してつくったプロトプラスト (protoplast) は分裂を繰り返して植物体を再生することができる。
陸上植物以外の植物 (藻類) では細胞外皮の構造は極めて多様である。細胞壁を欠くものや鱗片 (有機質、珪酸質、石灰など) で覆われるもの、珪酸質やβ-1,3-キシラン、β-1,4-マンナン、ペプチドグリカンなどできた細胞壁をもつものがある。またセルロース性の細胞壁をもつものも多いが、合成酵素複合体の形態には多様性が見られる。
細胞壁について
| キシログルカン (xyloglucan) | |
| β-1,3-β-1,4-グルカン | |
| グルクロノアラビノキシラン (glucuronoarabinoxylan) | |
| キシラン (xylan) =アラビノキシラン (arabinoxylan ) とグルクロノキシラン (glucuronoxylan) | |
| グルコマンナン (glucomannan) | |
| ガラクトマンナン (galactomannan) | |
| アラビノガラクタンII (arabinogalactan II) | |
| ホモガラクツロナン (homogalacturonan, HG) | |
| ラムノガラクツロナン I (rhamnogalacturonan I, RG-I) | |
| ラムノガラクツロナン II (rhamnogalacturonan II, RG-II) | |
| アピオガラクツロナン (apiogalacturonan) | |
| アラビノガラクタン (arabinogalactan) | |
| アラビナン (arabinan) | |
| ガラクタン (galactan) |
 細胞の生物学
細胞の生物学

