|
�s���t�ɂ���ׁA���iroot�j�̓����\���͑S�Ă̈ۊǑ��A����ʂ��đ��l�������Ȃ��B����́A�����ӂ��n���Ƃ����ω������Ȃ������ɂ���A���Ɩ��@�{���̋z���E�ʓ��ƐA���̂̎x���Ƃ����قƂ�ǂ̗���A���ŋ��ʂ̓�����S���Ă��邽�߂��ƍl������B
���̍ŊO�w���\���ŕ����Ă���A���̓����ɂ���w������B��w�̍œ��w�ɂ�����������A�ؕ���⿕����܂����S�����͂�ł���B���̐�[�ɂ������ŕی삳�ꂽ���[�����g�D������A���͂����Ő������Ă���B
|
�}�P. ��ʓI�ȍ��̓����\��
|
 ���� ����
���͂ӂ��n���ɂ���A��ɓy�Ɛ킢�Ȃ���L�����Ă���B�y���q�Ƃ̖��C���獪�̕����g�D��ی삷�邽�߁A���̐�[�������iroot cap�j�Ƃ������ȑg�D�ŕ����Ă���A���[�����g�D������Ă���B�����͑��זE�w����Ȃ�X�q��̍\���ŁA���[�����g�D����O���i��[���j�Ɍ����Č`�������B�����L������ɂ�č����זE�͊O������͂��ꗎ���A��������V�����זE���⋋�����B�����̍זE�͎��X�ƐV������Ă���A�`������Ă���͂��ꗎ����܂Ő������x�ł���B
�����̍זE�̓f���v�������S�t���i���V���[�Q�@mucilage�A�S���Q���@mucigel�j���������A�S���W���o�R�ő��ʂɕ��債�Ă���B�S�t���̓K���N�c�����_�d���̂��听���Ƃ��A�����m�|�X��t�R�[�X�Ȃǂ��܂܂��_�������ł���B�S�t���̑��݂ɂ���ēy�뗱�q�Ƃ̖��C���������č�����ی삷��ƂƂ��ɁA����L�����₷�����Ă���B���ۂɍd���y��ł͔S�t���̕���ʂ���������B���������y��ł́A�S�t���Ɠy��̍������ɂ���č��̎���ɏ���`�����A���̕ې��͂����߂Ă���B
�܂����̔S�t���́A���̎��́i�����@rhizosphaere�j�Ő����鐶���i�ۂ�ۗށj�ɐ���������Ă���B
���͂ӂ��d�͂̕��L�т����n���i�d�͋����j�������B���̎d�g�݂̒��ŁA�����̒�����ɑ��݂������t�זE�Ɋ܂܂���A�~���v���X�g���d�v�Ȗ������ʂ����Ă���炵���B
|
|
���[�����g�D
���̐�[�ɂ����[�����g�D�iroot apical meristem�j������A�����ŕی삳��Ă���B���[�����g�D����͍����ƍ����\���E��w�E���S����������B
�V���[�g�������g�D�Ɠ��l�A���[�����g�D�̍\���ɂ͑��l����������B�V�_�j�Ȃǂł́A���̃V���[�g���Ɠ��l�A�����P���n���זE���獪�̑S�g�D���`�������B���̎n���זE�����[�זE�Ƃ��l�ʑ̂�悵�A���ʂ���͍������A���ʂ���͑��̑g�D���`�������B�q�J�Q�m�J�Y���j�◇�q�A���ł́A�����̎n���זE�������g�D���`�����A��������S�g�D��������B��q�A���̍��[�����g�D�ɂ͈ȉ��ɋL���悤�ɕ��^�ƊJ���^���m���Ă���B
- ���^�iclosed type�j
- �����g�D�����Ăȑw��i�ӂ��R�w�j�ŁA�������獪���E�\��E��w�E���S�����`�������B�g�D�̗R���W����r�I���Ăł���B�����ƕ����g�D�͖��Ăɋ�ʂł���B�g�E�����R�V�i�C�l�ȁj�ȂǂɌ�����B
- �J���^�iopen type�j
- �����g�D�̑w��\�����s���ĂŁA�c�̍זE��ł����R�������icolumella�j���`������B�g�D�̗R���W�͕s���āB�����ƕ����g�D�̋�ʂ͂͂����肵�Ȃ��B�^�}�l�M�i�l�M�ȁj��\���}���i�}���ȁj�ȂǂɌ�����B
������������̍��̌���͑S�ĕ��^�ł��邪�A����L�����Ă����ɏ]���ĕ��^��ێ�������̂ƁA�n���זE�Q�������̕����։����o����ĊJ���^�ɕω�������̂ɕ������B
���[�����g�D�̒����ɂ́A�����̒Ⴂ�Î~���S�iquiescent center�j������B�Î~���S�͍��̑g�D�`�����R���g���[�����Ă���ƍl�����Ă���B�Î~���S�̂�������̕����A�O�����g�D�ipromeristem�j�ōł������ɍזE���s���Ă���A��w�⒆�S����������B�Î~���S�̐�[���i���ʁj�ɂ������n���זE�Q������A�����̍זE���`������Ă���B���̕\��͑O�����g�D�����������̂ƍ����n���זE�Q�����������̂�����B
- �����
- �זE������s���Ă���̈�B���[�����g�D�̂���̈�ł���A�זE�͏������B
- �L����
- �זE���L���������s���Ă���̈�B�L����̏㕔�t�߂ł͍��сi��q�j���ł������`�������B
- ���n��
- �זE�̕���E�L�����I�������̈�B�����ł͍��т��������ċz���т��`�����Ă���B�����E�L����ɂ���ׂ�ƃN�`�N���w�������Ȃ�B
|
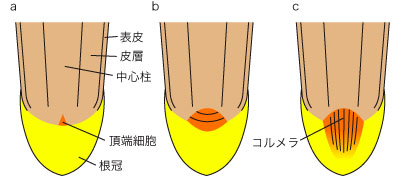
�}�R. ���[�����g�D�̍\���Ɍ����鑽�l��
a. �����P�̒��[�זE��������. b. ���^. c. �J���^.
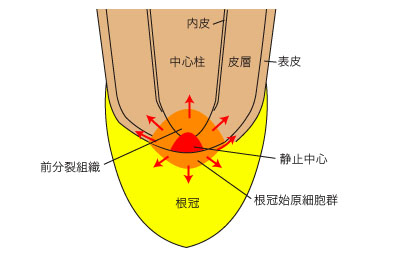
�}�S. ���[�����g�D
|
�\��
���̊튯�Ɠ��l�A�����\���iepidermis�j���ӂ��P�w���\��זE�iepidermal cell�j����Ȃ�B�܂��\��̓���̕�������͍��т��`�������i��q�Q���j�B���̕\��͂ӂ��n���ɂ��邽�߁A�N�`�N���w�̔��B�͎キ�A�܂��C�E���Ȃ��B
���̕\��͍����n���ŐL�����Ă����Ԃɂ͂��ꗎ���Ă��܂��B���̏ꍇ�A��������A���ł������iperiderm�j�A����ȊO�ł��O���iexodermis�j��������ی삵�Ă���B����̓R���N�`���w��������A�O��͔�w�̍ŊO�w���؉��E�����������̂ł���B
�����A���ł��郉���Ȃ�T�g�C���Ȃ̋C���̐�[���ł́A�\�炪���w�ɂȂ邱�Ƃ�����i���w�\���@multiple epidermis�j�A�זE�Ԍ����Đ������z���E�������Ă���B���̂悤�ȑg�D�������ivelamen�j�Ƃ����B
|
|
����
���̕\��̈ꕔ����́A�O���������ē�����я�ˋN��������B���̖я�ˋN�������iroot hair�j�Ƃ����A�y���q�̊Ԃɕ��������čג����L�т�B���т̒��a�͖�10 µm�قǂŁA�����͂��܂��܂ł���B���т͕\��זE���s�����Ăł����������זE�݂̂��琶����ꍇ������A���̍זE�����ь`���זE�i�����с@trichoblast�j�Ƃ����B
���т͍��Ɠy��Ƃ̐ڐG�ʂ傳���A����n���ɂȂ��~�߂铭�����ʂ����Ă���B�܂���ʂɂ͐��Ɩ��@�{���̋z���ɂ��d�v���Ƃ����邪�A���̋@�\�͍��ɋ�������ۗށi�ۍ��ہj���S���Ă��镔�����傫���i����A���̂قƂ�ǂ͋ۍ������j�A���ю��̂̊�^�ɂ����Ă͋^�⎋����ӌ�������B�����������̖��������܂�K�v�łȂ�����������ɂ͍��т͂��܂蔭�B���Ȃ��B
���т͍��̐L����Ō`������A�ӂ����̐L���ɔ����ď����͎����Ă����B���т̎����͂ӂ��������琔�T�Ԃł���B���������ɂ͔��N���琔�N�Ԑ������鍪�т�����A�h�������ipersistent root hair�j�Ƃ����B�h�����т̌`�ԂⒷ���͎���ٓI�ł���B
|
|
��w
��w�icortex�j�͕\��ƒ��S���̊Ԃɑ��݂���B���̔�w�͂����ނˍזE�ǂ������_�זE����Ȃ�A�f���v���Ȃǂ�����̂ɏd�v�Ȗ������ʂ����Ă���B�j���W���i�Z���ȁj��_�C�R���i�A�u���i�ȁj�ł͍��̔�w�������Ȃ��Ē����튯�ƂȂ��Ă���B�܂���w�ɂ͍זE�Ԍ��������A���̌ċz�̂��߂̃K�X�����ɖ𗧂��Ă���i��C�Ԍ��@air space�j�B���̔�w�`���ɂ�����זE����͋K���I�ł��邽�߁A�S�זE���P�̍זE�Ԍ������͂�ł��邱�Ƃ������B����͌s��t�Ɍ�����R�זE���͂ލזE�Ԍ������̐ς��傫���A��葽���̋�C�邱�Ƃ��ł���B
��w�̍ŊO�w�ɂ͂Ƃ����O���iexodermis�j������A�\��̂����ɍ���ی삷�������S���Ă���B�O��͑��݂ɖ��������P�w�i�Ƃ��ɐ��w�j�̍זE����Ȃ�A�זE�ǂ������X�x������O�j�����܂ށB�܂��������߂��Ւf����J�X�p���[���i��q�j�����݂��邱�Ƃ�����B
���̔�w�̍œ����ɂ͂ӂ����Ă������iendodermis�j������B����͂P�w�̍זE�w����Ȃ�A�זE�͑��݂ɖ������ĊԌ��͂Ȃ��B����Ɉꎟ�ǂɂ̓X�x��������Ȃ��J�X�p���[���iCasparian strip�j�������āA�זE�ǂ�ʂ����������߂��Ւf���Ă���B�܂�����זE�ɂ̓��O�j�����~�ς��邱�Ƃ�����B���̂��ߔ�w�ƒ��S���̊Ԃ̕��������́A�K������̍זE����ʉ߂��邱�ƂɂȂ�A����ɂ���Č����ʂ����߂���Ă���B�܂��זE�ǂ��������������̓��ߔ\���c���ʉߍזE�ipassage cell�j�����݂��邱�Ƃ�����B����͔����I�ɂ͔�w�̒��ōŌ�ɕ�������B
|
|
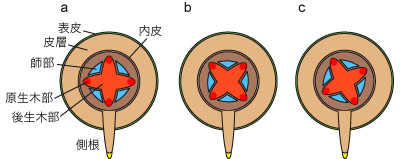
���S��
���ł́A���S�ɂ���ؕ������ˏ�ɘr��L���Ă���A���̘r�Ƙr�̊ԂɎt�������݂���B���̂悤�ȈۊǑ������ˈۊǑ��iradial vascular bundle�j�Ƃ�сA���̒��S�������˒��S���iactinostele�j�Ƃ����B�s�ł͂��܂��܂ȃ^�C�v�̒��S���������邪�A�S�Ă̈ۊǑ��A����ʂ��č��͕��˒��S���������Ă���B�P�q�t�A���ł́A���̒��S�����ؕ��ł͂Ȃ��A�_�g�D����Ȃ����ipith�j�ɂȂ��Ă��邱�Ƃ������B
���˒��S���ł́A�����ؕ����r�̐�[�i�O���j�ɂ���A�����t�����O���ɂ���B�܂荪�ł͖ؕ����t�����˂��O���^�iexoarch�j�ł���B
�����ؕ��̐��ɂ͈ȉ��̂悤�ȕψق�����B����͕��ތQ�ɂ���Ĉ��̌X���͂��邪�A����̓��ł��ψق��݂��邱�Ƃ�����B
- �ꌴ�^�imonoarch�j�F�����ؕ����P�B�~�Y�j�����ɂ݂���B
- �^�idiarch�j�F�����ؕ����Q�B�V�_�A���ɂ݂���B
- �O���^�itriarch�j�F�����ؕ����R�B�V�_�A���A���q�A���A�o�q�t�ނɂ݂���B
- �l���^�itetrarch�j�F�����ؕ����S�B���q�A���A�o�q�t�ނɂ݂���B
- �܌��^�ipentarch�j�F�����ؕ����T�B�o�q�t�ނɂ݂���B
- �����^�ipolyarch�j�F�����ؕ����P�B�����ؕ����U�ȏ�B�P�q�t�ނɂ݂���B
���S���̍ŊO�w�A�܂��w�œ��w�ł������̂��������ɂ͂P�w�̍זE�w����Ȃ������ipericycle�j������B����͕����g�D�̐�����ێ����Ă���A�Ⴂ���ł͂Ƃ���ǂ���Ő��w���N����A�܂������̌����������i���L�Q���j�B�Â����ł͓���̍זE�����\�͎����邪�A�������鍪�ł͓��₩��R���N�`���w��������i���L�Q���j�B
���S���̑��̕����͏_�g�D����Ȃ�B���S���̏_�g�D�ɂ͍זE�Ԍ����قƂ�ǂȂ��_�Ŕ�w�̏_�g�D�ƑΏƓI�ł���B
|
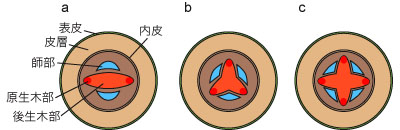
�}�W. �����ؕ��̐��ɂ�鍪�̃^�C�v
a. �^. b. �O���^. c. �l���^.
|
���̕��}
�������}����ɂ́A�s�Ɠ��l�A�厲�Ƒ����̋�ʂȂ��ɕ���������}�ƁA�厲���瑤�����ł��P�����}������B���̓��}�̓q�J�Q�m�J�Y���ނɌ����A�����g�D�����Ă��ꂼ�ꂪ�V���ȕ����g�D�ƂȂ�A�}�����ꂪ�N����B
�قƂ�ǂ̈ۊǑ��A���ł͒P�����}��������B�卪�̒��[�����◣�ꂽ�����������i���S���̍ŊO�w�j���V���ȕ����g�D�ƂȂ�A�卪�̔�w�ƕ\���˂��j���������ilateral root�j���L�т�B���̂悤�ɍ��̕��}�������I�iendogenous�j�ł���A�V���[�g�̂��ꂪ�O���I�iexogenous�j�ł���̂Ƒ傫���قȂ�B
�卪���瑤�����ł�ꏊ�́A��ɂ���Ăقڈ��ł���A�O�����猩��Ɖ��{���̏c����ɕ��z����B�����͎卪�̕��˒��S���ɂ����錴���ؕ��i���ˏ�̖ؕ��̘r�̐�[�j�̊O���A�܂��͎t���̊O������ł邱�Ƃ������A���̂��ߎ卪��̑����̏c���猴���ؕ��̐�������ł���B�Ⴆ�_�C�R���i�A�u���i�ȁj�ł͓^�A�j���W���i�Z���ȁj�͎l���^�A�T�c�}�C���i�q���K�I�ȁj�͌܌��^�ł��邱�Ƃ��킩��B�����������^�̍��̏ꍇ�A�����̈ۊǑ��������ɐN������ꍇ������A�����ؕ��̐��𐄒肷��͍̂���ł���B
|
�}�X. �����̔�������ʒu
a. �����ؕ��̊O��. b. �t���̊O��. c. �ؕ��Ǝt���̊�.
|
�s�荪�̔���
�c������т��̑����ȊO���琶�������̂��Ƃ��s�荪�iadventitious root�j�Ƃ����B�s�荪�͈ۊǑ��A���ɂ����ċɂ߂Ăӂ��Ɍ����A���ɒP�q�t�A���ł͍��̂قƂ�ǂ��s�荪�ł���B
���̕��}�������I�Ȃ̂Ɠ��l�A�s�荪�̔������قƂ�ǂ������I�ł���A�ӂ��s�̐߂ɂ����������ɑ������镔�����琶����B�܂�Ƀx�S�j�A�i�V���E�J�C�h�E�ȁj�̗t�}���ɂ����ėt�̕\�ʂ���s�荪��������悤�ɁA�O���I�ɕs�荪�������邱�Ƃ�����B
|
|
���̓���
�P�q�t�A���ł́A�s�Ɠ��������ł������͔��ɂ܂�ł���B����ɑ��đo�q�t�A���ł́A���͂ӂ������i��听���j���s���A���ɖؖ{�ł͐A���̂̎x���Ƃ����Ӗ��ŏd�v�ł���B
���̈ꎟ�ؕ��ƈꎟ�t���̊Ԃ��ۊǑ��`���w�ivascular cambium�j�������A�זE������ɍs����悤�ɂȂ�B���̌��ʂƂ��ĉ��f�ʂŐ��`�����Ă����ؕ��͂����Ɋۂ��Ȃ�B�s�Ɠ��l�A�ۊǑ��`���w�͓������ؕ����A�O�����t�����`������B
������听�����Ă����ƁA���Ƃ��Ƃ����������ꎟ��w�A�\��͉��Ă��܂��B�����ɁA���S���̍ŊO�w�ł������������R���N�`���w�icork cambium, phellogen�j���h�����A�������`�������B����͕\��̂����ɍ���ی삷��@�\���ʂ����悤�ɂȂ�B
|
|


