|
茎や葉にくらべ、根 (root) の内部構造は全ての維管束植物を通じて多様性が少ない。これは、根がふつう地中という変化が少ない環境下にあり、水と無機養分の吸収・通道と植物体の支持というほとんどの陸上植物で共通の働きを担っているためだと考えられる。
根の最外層は表皮で覆われており、その内側には皮層がある。皮層の最内層には内皮があり、木部と篩部を含む中心柱を囲んでいる。根の先端には根冠で保護された根端分裂組織があり、根はここで成長している。
|
図1. 一般的な根の内部構造
|
 根冠 根冠
根はふつう地中にあり、常に土と戦いながら伸長している。土粒子との摩擦から根の分裂組織 (根端分裂組織) を保護するため、根の先端は根冠 (root cap) とよばれる特殊な組織で覆われている。根冠は多細胞層からなる帽子状の構造で、根端分裂組織から外側 (先端側) に向けて形成される。根が伸長するにつれて根冠細胞は外側からはがれ落ち、内側から新しい細胞が補給される。根冠の細胞は次々と新生されており、形成されてからはがれ落ちるまで数日程度である。
根冠の細胞はデンプンから粘液質 (ムシラーゲ mucilage、粘質ゲル mucigel) を合成し、ゴルジ体経由で多量に分泌している。粘液質はガラクツロン酸重合体を主成分とし、ラムノ-スやフコースなどが含まれる酸性多糖である。粘液質の存在によって土壌粒子との摩擦を減少して根冠を保護するとともに、根を伸長しやすくしている。実際に硬い土壌では粘液質の分泌量が増加する。乾燥した土壌では、粘液質と土壌の混合物によって根の周りに鞘を形成し、根の保水力を高めている。
またこの粘液質は、根の周囲 (根圏 rhizosphaere) で生きる生物 (細菌や菌類) に生活環境を提供している。
根はふつう重力の方へ伸びる屈地性 (重力屈性) を示す。この仕組みの中で、根冠の中央基部に存在する平衡細胞に含まれるアミロプラストが重要な役割を果たしているらしい。
マキ (マキ科) やトチノキ (トチノキ科)、トキソウ (ラン科) などでは根冠を欠く根が存在する。
また一般的な根の機能をもたない水生植物のウキクサ (サトイモ科)、トチカガミ (トチカガミ科)、ホテイアオイ (ミズアオイ科) など、寄生植物のスナヅル (クスノキ科)、ネナシカズラ (ヒルガオ科)、ハマウツボ (ハマウツボ科) など、腐生植物のギンリョウソウやイチヤクソウ (ツツジ科) などでは根冠が未発達である。上記の内、ウキクサやホテイアオイなど水生植物では不定根が出るときに母器官の組織が根端に取り残されて根冠状の組織になり、この組織は根帽 (根嚢 root pocket) とよばれる。通常の根冠をもつアカウキクサ (ウラボシ綱) やセキショウモ (トチカガミ科) にも根帽が見られる。
|
|
根端分裂組織
根の先端には根端分裂組織 (root apical meristem) があり、根冠で保護されている。根端分裂組織からは根冠と根の表皮・皮層・中心柱がつくられる。
シュート頂分裂組織と同様、根端分裂組織の構造には多様性が見られる。シダ綱などでは、そのシュート頂と同様、ただ1個の始原細胞から根の全組織が形成される。この始原細胞は頂端細胞とよばれ四面体を呈し、下面からは根冠が、側面からは他の組織が形成される。ヒカゲノカズラ綱や裸子植物では、複数の始原細胞が分裂組織を形成し、そこから全組織がつくられる。被子植物の根端分裂組織には以下に記すように閉鎖型と開放型が知られている。
- 閉鎖型 (closed type)
- 分裂組織が明瞭な層状 (ふつう3層) で、そこから根冠・表皮・皮層・中心柱が形成される。組織の由来関係が比較的明瞭である。根冠と分裂組織は明瞭に区別できる。トウモロコシ (イネ科) などに見られる。
- 開放型 (open type)
- 分裂組織の層状構造が不明瞭で、縦の細胞列であるコルメラ (columella) を形成する。組織の由来関係は不明瞭。根冠と分裂組織の区別ははっきりしない。タマネギ (ネギ科) やソラマメ (マメ科) などに見られる。
生成したばかりの根の原基は全て閉鎖型であるが、原基が伸長していくに従って閉鎖型を保持するものと、始原細胞群が根冠の方向へ押し出されて開放型に変化するものに分かれる。
根端分裂組織の中央には、分裂活性の低い静止中心 (quiescent center) がある。静止中心は根の組織形成をコントロールしていると考えられている。静止中心のすぐ基部側の部分、前分裂組織 (promeristem) で最も活発に細胞分裂が行われており、皮層や中心柱がつくられる。静止中心の先端側 (遠位) には根冠始原細胞群があり、根冠の細胞が形成されている。根の表皮は前分裂組織からつくられるものと根冠始原細胞群からつくられるものがある。
- 分裂域
- 細胞分裂を行っている領域。根端分裂組織のある領域であり、細胞は小さい。
- 伸長域
- 細胞が伸長成長を行っている領域。伸長域の上部付近では根毛 (後述) が最も多く形成される。
- 成熟域
- 細胞の分裂・伸長が終了した領域。下部では根毛が密生して吸水帯を形成している。分裂域・伸長域にくらべるとクチクラ層が厚くなる。
|
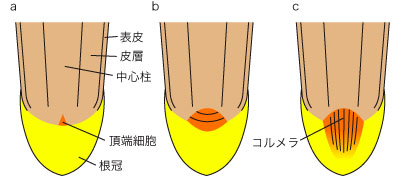
図3. 根端分裂組織の構造に見られる多様性
a. ただ1つの頂端細胞をもつもの. b. 閉鎖型. c. 開放型.
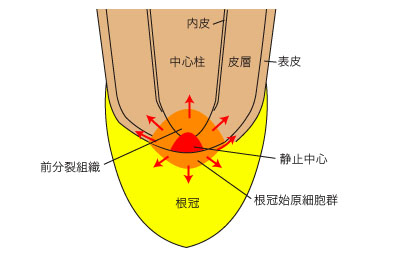
図4. 根端分裂組織
|
表皮
他の器官と同様、根の表皮 (epidermis) もふつう1層の表皮細胞 (epidermal cell) からなる。また表皮の特定の部分からは根毛が形成される (後述参照)。根の表皮はふつう地中にあるため、クチクラ層の発達は弱く、また気孔もない。
根の表皮は根が地中で伸長していく間にはがれ落ちてしまう。この場合、二次成長する植物では周皮 (periderm)、それ以外では外皮 (exodermis) が内部を保護している。周皮はコルク形成層からつくられ、外皮は皮層の最外層が木化・厚化したものである。
着生植物であるラン科やサトイモ科の気根の先端部では、表皮が多層になることがあり (多層表皮 multiple epidermis)、細胞間隙を生じて水分を吸収・貯蔵している。このような組織を根被 (velamen) という。
|
|
根毛
根の表皮の一部からは、外側へ向かって特殊な毛状突起が生じる。この毛状突起を根毛 (root hair) といい、土粒子の間に分け入って細長く伸びる。根毛の直径は約10 µmほどで、長さはさまざまである。根毛は表皮細胞が不等分裂してできた小さい細胞のみから生じる場合があり、この細胞を根毛形成細胞 (原根毛 trichoblast) という。
根毛は根と土壌との接触面を増大させ、根を地中につなぎ止める働きを果たしている。また一般には水と無機養分の吸収にも重要だといわれるが、この機構は根に共生する菌類 (菌根菌) が担っている部分が大きく (陸上植物のほとんどは菌根をもつ)、根毛自体の寄与についいては疑問視する意見もある。しかしこれらの役割があまり必要でない水中根や寄生根には根毛はあまり発達しない。
根毛は根の伸長域で形成され、ふつう根の伸長に伴って順次枯死していく。根毛の寿命はふつう数日から数週間である。しかし中には半年から数年間生存する根毛もあり、宿存根毛 (persistent root hair) とよばれる。宿存根毛の形態や長さは種特異的である。
|
|
皮層
皮層 (cortex) は表皮と中心柱の間に存在する。根の皮層はおおむね細胞壁が薄い柔細胞からなり、デンプンなどを貯蔵するのに重要な役割を果たしている。ニンジン (セリ科) やダイコン (アブラナ科) では根の皮層が厚くなって貯蔵器官となっている。また皮層には細胞間隙が多く、根の呼吸のためのガス交換に役立っている (空気間隙 air space)。根の皮層形成における細胞分裂は規則的であるため、4細胞が1つの細胞間隙を取り囲んでいることが多い。これは茎や葉に見られる3細胞が囲む細胞間隙よりも体積が大きく、より多くの空気を得ることができる。
皮層の最外層にはときに外皮 (exodermis) があり、表皮のかわりに根を保護する役割を担っている。外皮は相互に密着した1層 (ときに数層) の細胞からなり、細胞壁が厚くスベリンやリグニンを含む。また物質透過を遮断するカスパリー線 (後述) が存在することがある。
根の皮層の最内部にはふつう明瞭な内皮 (endodermis) がある。内皮は1層の細胞層からなり、細胞は相互に密着して間隙はない。さらに一次壁にはスベリンからなるカスパリー線 (Casparian strip) があって、細胞壁を通じた物質透過を遮断している。また内皮細胞にはリグニンが蓄積することもある。このため皮層と中心柱の間の物質交換は、必ず内皮の細胞内を通過することになり、これによって交換量が調節されている。また細胞壁が厚化せず物質の透過能を残す通過細胞 (passage cell) が存在することがある。内皮は発生的には皮層の中で最後に分化する。
|
|
中心柱
根では、中心にある木部が放射状に腕を伸ばしており、その腕と腕の間に師部が存在する。このような維管束を放射維管束 (radial vascular bundle) とよび、その中心柱を放射中心柱 (actinostele) という。茎ではさまざまなタイプの中心柱が見られるが、全ての維管束植物を通じて根は放射中心柱をもっている。単子葉植物では、根の中心部が木部ではなく、柔組織からなる髄 (pith) になっていることが多い。また特殊な例として、ラン科、ヤシ科、シムラニンジン () などでは1つの根に数個~十数個の放射中心柱をもつことがある。
放射中心柱では、原生木部が腕の先端 (外側) にあり、原生師部も外側にある。つまり根では木部も師部もつねに外原型 (exoarch) である。
原生木部の数には以下のような変異がある。これは分類群によって一定の傾向はあるが、同一個体内でも変異がみられることがある。
- 一原型 (monoarch) :原生木部が1つ。ミズニラ属などにみられる。
- 二原型 (diarch) :原生木部が2つ。シダ植物にみられる。
- 三原型 (triarch) :原生木部が3つ。シダ植物、裸子植物、双子葉類にみられる。
- 四原型 (tetrarch) :原生木部が4つ。裸子植物、双子葉類にみられる。
- 五原型 (pentarch) :原生木部が5つ。双子葉類にみられる。
- 多原型 (polyarch) :原生木部が1つ。原生木部が6以上。単子葉類にみられる。
中心柱の最外層、つまり皮層最内層である内皮のすぐ内側には1層の細胞層からなる内鞘 (pericycle) がある。内鞘は分裂組織の性質を保持しており、若い根ではところどころで垂層分裂が起こり、また側根の原基が発生する (下記参照)。古い根では内鞘の細胞分裂能は失われるが、二次成長する根では内鞘からコルク形成層が生じる (下記参照)。
中心柱の他の部分は柔組織からなる。中心柱の柔組織には細胞間隙がほとんどない点で皮層の柔組織と対照的である。
|
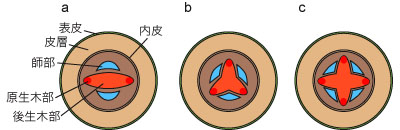
図8. 原生木部の数による根のタイプ
a. 二原型. b. 三原型. c. 四原型.
|
根の分枝
根が分枝するには、茎と同様、主軸と側軸の区別なく二又に分かれる二又分枝と、主軸から側軸がでる単軸分枝がある。
根の二又分枝は、茎の二又分枝と同様、ヒカゲノカズラ類に見られ、分裂組織が二分してそれぞれが新たな分裂組織となり、枝分かれが起きる。
ほとんどの維管束植物では単軸分枝が見られる。主根の頂端からやや離れた部分の内鞘 (中心柱の最外層) が新たな分裂組織となり、主根の皮層と表皮を突き破って側根 (lateral root) が伸びる。このように根の分枝は内生的 (endogenous) であり、シュートのそれが外生的 (exogenous) であるのと大きく異なる。
主根から側根がでる場所は、種によってほぼ一定であり、外側から見ると何本かの縦軸上に分布する。側根は主根の放射中心柱における原生木部 (放射状の木部の腕の先端) の外側、または師部の外側からでることが多く、そのため主根上の側根の縦列数から原生木部の数が推定できる。例えばダイコン (アブラナ科) では二原型、ニンジン (セリ科) は四原型、サツマイモ (ヒルガオ科) は五原型であることがわかる。しかし多原型の根の場合、複数の維管束が側根に侵入する場合があり、原生木部の数を推定するのは困難である。
|
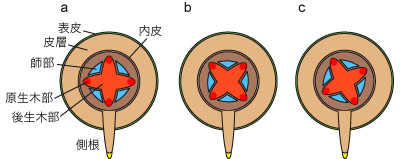
図9. 側根の発生する位置
a. 原生木部の外側. b. 師部の外側. c. 木部と師部の間.
|
不定根の発生
幼根およびその側根以外から生じた根のことを不定根 (adventitious root) という。不定根は維管束植物において極めてふつうに見られ、特に単子葉植物では根のほとんどが不定根である。
根の分枝が内生的なのと同様、不定根の発生もほとんどが内生的であり、ふつう茎の節における内鞘に相当する部分から生じる。まれにベゴニア (シュウカイドウ科) の葉挿しにおいて葉の表面から不定根が生じるように、外生的に不定根が生じることがある。
|
|
根の二次成長
単子葉植物では、茎と同じく根でも二次成長は非常にまれである。それに対して双子葉植物では、根はふつう二次成長 (肥大成長) を行い、特に木本では植物体の支持という意味で重要である。
根の一次木部と一次師部の間に維管束形成層 (vascular cambium) が生じ、細胞分裂が盛んに行われるようになる。その結果として横断面で星形をしていた木部はすぐに丸くなる。茎と同様、維管束形成層は内側に二次木部を、外側に二次師部を形成する。
根が肥大成長していくと、もともとあった内皮や一次皮層、表皮は壊れてしまう。かわりに、中心柱の最外層である内鞘からコルク形成層 (cork cambium, phellogen) が派生し、周皮が形成される。周皮は表皮のかわりに根を保護する機能を果たすようになる。
|
|


