
![]()
![]()
![]()
![]()
![]()
| ユーグレノゾア (EUGLENOZOA): ねむり病原虫とユーグレナをふくむ初期真核生物 |
| ユーグレナ植物 画像データ | |
| 独立栄養性ユーグレナ類 | |
| 従属栄養性ユーグレナ類 | |
| キネトプラスト類(ボド類) |
ゾウリムシとならんで,なぜかミドリムシ(Euglena)は子供から大人までほとんどの日本人が知っている数少ない単細胞生物で,水中の代表的な生物として定着しています。しかし,実際にはこれほど理解が困難な生物はそうはありません。
ミドリムシは葉緑体をもち,光合成を行って生活しています。その点では「植物」といえます。しかし一方で,ミドリムシはユーグレナ運動 (euglenoid movement) とよばれる,細胞を頻繁に変形する運動を行い,また鞭毛を用いて遊泳します。ミドリムシはいわば植物でも動物でもない,不思議な生物です。多くの生徒や学生がミドリムシの分類上の位置に疑問をかかえたまま卒業しているのが現状でしょう。これは生物を動物と植物に二分する分類法が現在も一般的であるからです。
海綿から哺乳類まで,われわれの思い浮かべる動物は大変まとまりのある仲間です。つまり,すべての動物はたった一つの祖先生物から分化してきたものと考えられ,単系統群として認識されます。しかし,植物(光合成生物)はそうではありません。「植物」は光合成という機能をもつことでまとめられた生物群です。しかし,葉緑体の獲得が複数の細胞共生に由来することが理解された現在では,「光合成」は単一の系統群を認める根拠とはなり得ません。光合成生物は起源の異なる多数の系統群を含んでいます。ユーグレナはそのひとつで,実際には真核生物全体の進化の中で興味深い位置を占めています。
ユーグレナ類はおよそ40属800種を含む真核光合成生物の一群で,その大部分が単細胞遊泳性の鞭毛生物です。Eutreptia, Euglena, Trachelomonas, Phacusのように葉緑体をもつなかまのほか,吸収(Astasia)や食作用による従属栄養(Peranema, Entosiphon)で生息するものまであり,形態と生態は多様性に富んでいます。
ユーグレナ植物は次のような形態的特徴をもっています。
| ユーグレナ類の細胞外被はペリクル(pellicle)とよばれる。細胞膜の内側に蛋白質性のリボン状の裏打ち構造 (pellicle strip)を形成し,多数のstripが細胞をらせん状に取り巻いている。pellicle stripは微小管,小胞体,そしてアクチン,ミオシンを伴っており,隣り合うペリクル同士の滑りによりユーグレナ運動を引き起こす。 |
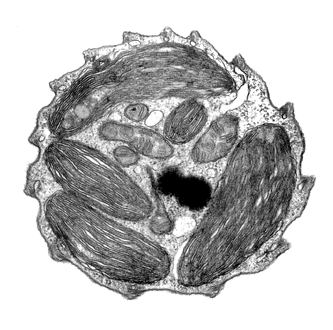 | |||
ミトコンドリアクリステは根元がくびれたうちわ形(ディスク形)である。 板状クリステ (flat cristae)および管状クリステ
(tubular cristae)と比較してください。 |
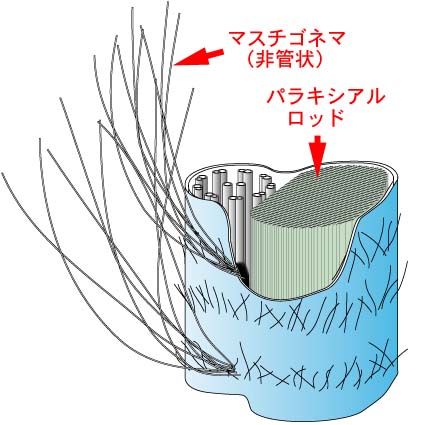
鞭毛はいわゆる片羽型といわれるもので,全面を覆う短いフェルト状の毛の他に微細な小毛が1列らせん状に取り巻いている。 この小毛は黄色植物で知られる管状マスチゴネマとは異なり,管状ではない。 |
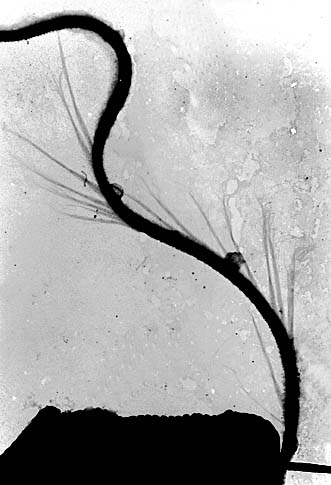 |
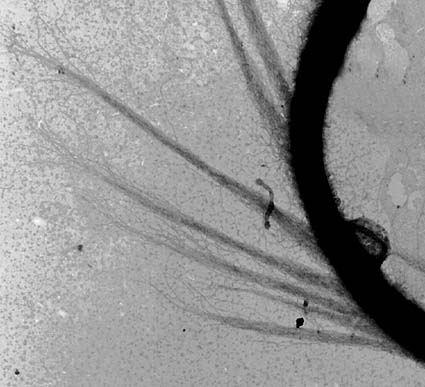 |
| 軸糸に沿ってparaxial rodが存在する。ユーグレナ植物とキネトプラスチダ類に特有。 | 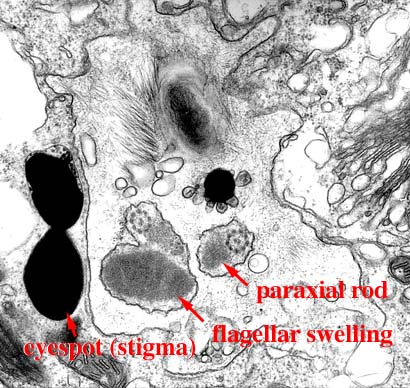 |
 | ||
鞭毛装置は3つの微小管性鞭毛根をもつ(dorsal root,
ventral root, intermediate root)。 これらの鞭毛根はキネトプラスチダ類のものと相同と考えられています。 |
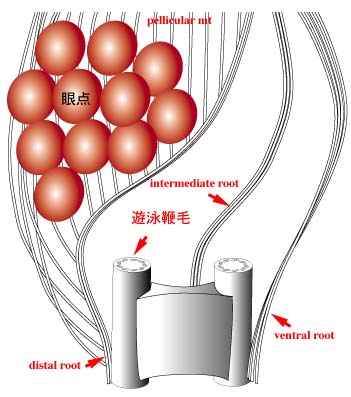 |
眼点は葉緑体の外,鞭毛溝の側部に位置し,長鞭毛の膨潤部とともに光受容装置を構成する。眼点は光の遮蔽あるいは集光の役割を果たしていると考えられる。 鞭毛膨潤部と眼点の関係は黄色植物と類似しています。 |
|
葉緑体は3枚の包膜に包まれる。 外側の包膜の起源についてはまだはっきりしたことは分かっていませんが,共生体(真核藻類)の細胞膜か,共生体が取り込まれたときの食胞の膜のどちらかに由来すると考えられます。 |
| 中間期の核でも染色体は凝集して存在する。 |
核分裂は完全に閉鎖型で核膜が残存し,紡錘体は核内に形成される。中期赤道板は形成されない。またしばしば分裂中も仁が消失せずに残る。 この性質もキネトプラスチダ類と類似しています。 |
葉緑体の共生起源
多くの真核光合成生物と同様に,ユーグレナ類の葉緑体も真核藻類の共生によって獲得されたと考えられています。現在まで共生体となった真核緑色藻類は特定されていませんが,緑色植物のアオサ藻やプラシノ藻が候補としてあげられています。
一方,宿主となった生物については,形態的特徴から鞭毛性原生生物キネトプラスト類
(Kinetoplastida)に近縁の生物であることがほぼ明らかにされています。これについてはユーグレノゾア (EUGLEMOZOA)の項目をみてください。
![]()
![]()
![]()
![]()
![]()