![]()
![]()
![]()
![]()
![]()

|
プラシノ藻綱(Prasinophyceae) |
|---|---|
|
アオサ藻綱(Ulvophyceae) |
|
緑藻綱(Chlorophyceae) |
|
シャジク藻綱(Charophyceae) |
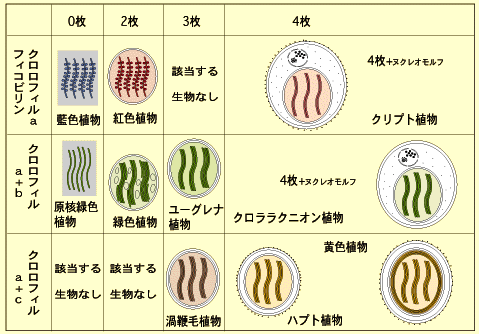
ミドリムシなどのユーグレナ植物やクロララクニオン植物もクロロフィルa・bをもちますが,細胞のつくりは著しく異なります。この相違は細胞共生によって説明されます。 | |
 葉緑体は2重膜に包まれ,他の包膜をもたない。 葉緑体は2重膜に包まれ,他の包膜をもたない。このような性質を持つ葉緑体は紅色植物と灰色植物で知られているだけで,他の藻類では葉緑体は2重膜の外に1枚または2枚の膜をもっています。 | |
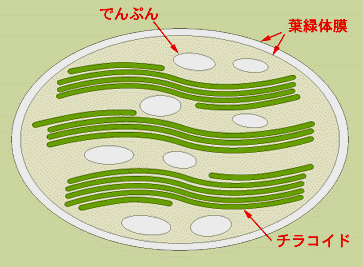 貯蔵物質としてでんぷん(starch)を葉緑体内部に形成する。 貯蔵物質としてでんぷん(starch)を葉緑体内部に形成する。他のすべての光合成生物は貯蔵物質を細胞質に蓄積しています。 | |
藻類には,他に管状クリステをもつ仲間とディスク状クリステをもつ仲間があります。 |
![]() UCMPの光合成色素のページ
(http://ucmp1.berkeley.edu/plants/pigments.html)
UCMPの光合成色素のページ
(http://ucmp1.berkeley.edu/plants/pigments.html)
緑色植物の葉緑体(シオグサ属) ![]()
光合成真核生物のなかで葉緑体が2枚の葉緑体膜だけをもち,その他の包膜をもたないのは緑色植物と紅色植物(紅藻類)および灰色植物だけです。灰色植物は藍藻が共生して葉緑体として定着する過程にある生物と考えられていますが,同様に緑色植物と紅色植物も1度だけの共生によって葉緑体が獲得されたと考えられています。これら3者の葉緑体が複数の共生によって互いに独立に獲得されたか,あるいは1度だけの共生で獲得されたのか現在でも異論のあるところです(細胞共生のページを参照)。しかし,もし葉緑体の起源が1つならば,灰色植物,紅色植物,緑色植物は単系統ということになり,これらを「植物界(Kingdom Planta)」としてまとめることが可能です。 Cavalier-Smithの8界説の植物界はこの考えに立ったものです。
緑色植物は一般に「緑藻」とよばれる藻類を含む仲間ですが,「緑藻」は現在では複数の大きな系統をふくんでいることが分かっており,新たな分類が提唱されています。これは細胞構造と細胞分裂の微細構造の比較形態学の成果に基づいていますが,分子系統の結果も大筋において微細構造から想定された分類系を支持しています。
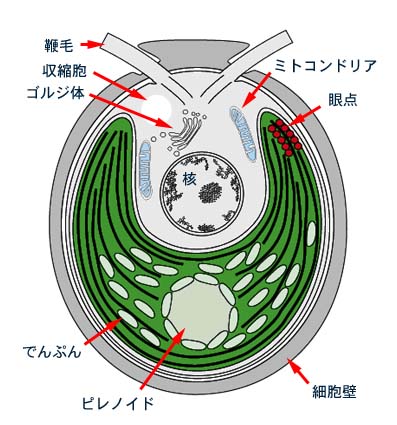 藻類の分類群は葉緑体の性質とともに,遊泳細胞(生殖細胞や単細胞遊泳性の種類)のもつ形態で特徴づけられます。緑色植物の遊泳細胞は一般に次の特徴をもっています。
藻類の分類群は葉緑体の性質とともに,遊泳細胞(生殖細胞や単細胞遊泳性の種類)のもつ形態で特徴づけられます。緑色植物の遊泳細胞は一般に次の特徴をもっています。
| 鞭毛は等長でムチ型,通常細胞の先端から生じ,等長である。 | |
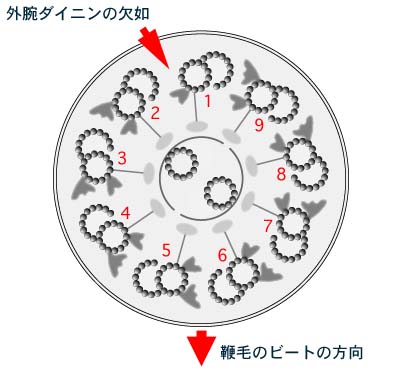 鞭毛軸糸のひとつは外腕のダイニン
(outer dynein) を欠く。ダイニンを欠くこの軸糸をNo.1と名付け,鞭毛先端からみて反時計回りに番号をふると,鞭毛のビート面はNo.5とNo.6の間を通る面になる。 鞭毛軸糸のひとつは外腕のダイニン
(outer dynein) を欠く。ダイニンを欠くこの軸糸をNo.1と名付け,鞭毛先端からみて反時計回りに番号をふると,鞭毛のビート面はNo.5とNo.6の間を通る面になる。 | |
 鞭毛移行部 (flagellar
transition region) に星状パターン構造 (stellate pattern) がある。 鞭毛移行部 (flagellar
transition region) に星状パターン構造 (stellate pattern) がある。 |
(注)これらの性質のうち,鞭毛の特徴はプラシノ藻綱にはあてはまらない(プラシノ藻綱のページを参照)。
緑色植物の分類を考える上で最も重要な細胞構造は鞭毛装置(flagellar apparatus)で,鞭毛と鞭毛の基部につながる基底小体(basal
body)およびこれに付随する繊維構造と微小管を含む複雑な構造です。
緑色植物は鞭毛の生じ具合から2つの異なるグループに分けることができます。
| 鞭毛は細胞前端から左右対称に生じる。このタイプの鞭毛は緑色の藻類の大部分に見られる。 | |
| 鞭毛は細胞の前端のやや後方から平行に生じ,側方または後方に向けて伸びる。このタイプの鞭毛はシャジク藻類のほか,淡水産の緑色藻類の一部(コレオケーテ(Coleochaete),キートスフェリディーム(Chaetosphaeridium)など)にみられる。 |
これら2つのグループは微細構造の点でも大きく異なっています。
![]() 左右対称の鞭毛をもつ仲間では,2つの基底小体それぞれの基部から2組の微小管が伸び,全体として十文字に見えます。この微小管は鞭毛根(flagellar
roots, flagellar rootlets)とよばれます。十字に見えるためにこのタイプの微小管は十字型鞭毛根系(cruciate
root system),または交叉するようにみえることから交叉型鞭毛根系とよばれます。4本の鞭毛根は細胞表面を後方に伸びます。
左右対称の鞭毛をもつ仲間では,2つの基底小体それぞれの基部から2組の微小管が伸び,全体として十文字に見えます。この微小管は鞭毛根(flagellar
roots, flagellar rootlets)とよばれます。十字に見えるためにこのタイプの微小管は十字型鞭毛根系(cruciate
root system),または交叉するようにみえることから交叉型鞭毛根系とよばれます。4本の鞭毛根は細胞表面を後方に伸びます。
もう少し詳しく見ると交叉型鞭毛根系は次のような特徴をもつことがわかります。
![]() 一つの基底小体から生じる鞭毛根は2種類あり,ひとつは数本(多くの場合2本)の微小管からなり,基底小体の基部を上に向けたとき,右下に伸びます。この鞭毛根は2-rootまたはright
rootとよばれます。もうひとつは複数の(ほとんどの場合4本以上)の微小管からなり,基底小体の基部を上に向けたとき,左下に伸びます。この鞭毛根はX-rootまたはleft rootとよばれます。
一つの基底小体から生じる鞭毛根は2種類あり,ひとつは数本(多くの場合2本)の微小管からなり,基底小体の基部を上に向けたとき,右下に伸びます。この鞭毛根は2-rootまたはright
rootとよばれます。もうひとつは複数の(ほとんどの場合4本以上)の微小管からなり,基底小体の基部を上に向けたとき,左下に伸びます。この鞭毛根はX-rootまたはleft rootとよばれます。
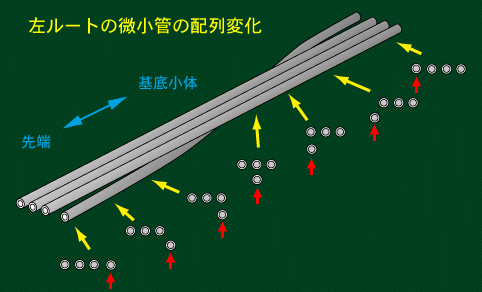 X-root (left root)は左側の1本が他の微小管の下にもぐり,やがて右側に出てくるという微小管の配列変化(configurational
change)を示すのが特徴です。
X-root (left root)は左側の1本が他の微小管の下にもぐり,やがて右側に出てくるという微小管の配列変化(configurational
change)を示すのが特徴です。
このような性質から交叉型鞭毛根系はX-2-X-2 root systemともよばれます。
交叉型鞭毛根系は中心を軸にして180度回転すると1組の基底小体と鞭毛根系が他方のそれに重なるという性質をもっています。これは180度回転対称とよばれます。
鞭毛が平行に生じるコレオケーテなどの遊泳細胞では,1個の基底小体だけが微小管性の鞭毛根をもっています。鞭毛根は多数の微小管(スプライン:splineとよばれる)からなり,その基部に層状の構造が付随するのが特徴です。スプラインと層状構造をあわせて多層構造体(multilayered
structure: MLS)とよびます。このスプライン微小管はright rootと相同であることが分かっています(プラシノ藻綱のページを参照)。このほかにleft
root にあたる数本の微小管からなる鞭毛根が1つ存在ます。

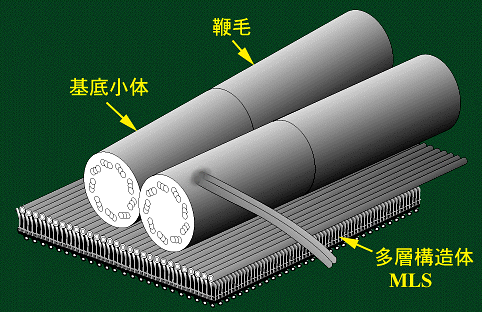
このように,これらの緑色藻類の鞭毛装置は著しく非対称の構造をもっており,交叉型鞭毛装置と明瞭に区別されます。
MLSはコケ,シダおよび裸子植物(イチョウ,ソテツ類)の精子にも存在します。このことは,緑色藻類のなかには陸上植物に似た仲間とそうでないものが存在していること,すなわち緑色藻類は複数の系統群を含んでいることを示唆しています。
交叉型鞭毛根は緑色藻類に広く分布していますが,詳細に観察すると,さらに基底小体の配列が異なる2つのタイプに分かれることが分かります。これらは基底小体の「ずれ(offset)の違いにより,時計回り配列(clockwise orientation=CW)(または1時-7時配列(1 o'clock-7 o'clock (1/7) orientation))と反時計回り配列(counterclockwise orientation=CCW)(または11時-5時配列(11 o'clock-5 o'clock (11/5) orientation))とよばれます。
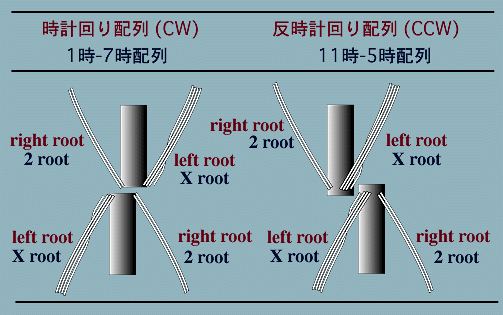 これら2つの交叉型鞭毛装置は基底小体の基部の配列でも区別できます。時計回り配列をもつ鞭毛装置では,2つの基底小体は基部を向かい合わせて配列し,基底小体のほぼ半分ほど時計回りにずれています。そして基底小体は決して重なり合うことはありません。これに対して反時計回り配列の鞭毛装置では,基底小体の基部が例外なく重なり合うのが特徴です。
これら2つの交叉型鞭毛装置は基底小体の基部の配列でも区別できます。時計回り配列をもつ鞭毛装置では,2つの基底小体は基部を向かい合わせて配列し,基底小体のほぼ半分ほど時計回りにずれています。そして基底小体は決して重なり合うことはありません。これに対して反時計回り配列の鞭毛装置では,基底小体の基部が例外なく重なり合うのが特徴です。
![]() 時計回り配列(CW)の交叉型鞭毛装置は大部分の淡水産の緑色藻類にみられます。これに対して,反時計回り配列(CCW)の鞭毛装置はほとんどの海産の緑色植物(つまり緑色の海藻のほとんど)の遊走子や精子にみられます。例外はありますが,一般に淡水産と海産の緑色藻類は,鞭毛の生じる様式においても違いがあります。クラミドモナスなどの多くの淡水産の緑色藻類では基底小体が約90度に開き,V字型に生じます。これに対して海産の緑色藻類では基底小体はほぼ水平方向に生じているのが普通です。
時計回り配列(CW)の交叉型鞭毛装置は大部分の淡水産の緑色藻類にみられます。これに対して,反時計回り配列(CCW)の鞭毛装置はほとんどの海産の緑色植物(つまり緑色の海藻のほとんど)の遊走子や精子にみられます。例外はありますが,一般に淡水産と海産の緑色藻類は,鞭毛の生じる様式においても違いがあります。クラミドモナスなどの多くの淡水産の緑色藻類では基底小体が約90度に開き,V字型に生じます。これに対して海産の緑色藻類では基底小体はほぼ水平方向に生じているのが普通です。
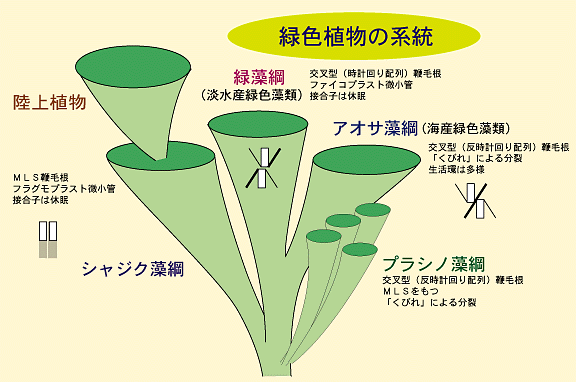
このように,鞭毛装置からみると,緑色植物は3つの大きな仲間に分けられます。
MLS型鞭毛装置をもつ,陸上植物を含むグループ
である。
鞭毛装置の微細構造と平行して研究されたのが微細構造のレベルでの細胞分裂の様式です。緑色植物には,核分裂の様式や紡錘体の挙動,あるいは分裂の最終段階で核と細胞質を二分するしくみなどの性質が異なる細胞分裂が存在し,それら異なる細胞分裂の様式の分布は鞭毛装置に基づく分類にほぼ一致しています。
よく知られているように,陸上植物では核の分裂時に核膜が消失します。つまり分裂の中期から終期にかけて染色体は細胞質中にむき出しになっています。終期になると極紡錘体の伸長によって染色体が両極に移動して娘核が形成され,やがて細胞板(cell plate)が形成されます。このとき最後まで残っていた中間紡錘体(persistent interzonal spindle)が染色体から離れて赤道面に沿って分裂軸と平行に並ぶようになります。細胞板形成時に出現するこの構造は古く光学顕微鏡で観察され,フラグモプラスト(隔膜形成体,phragmoplast)と呼ばれていたもので,フラグモプラスト微小管とよばれます。細胞板は小胞(ゴルジ体に由来する)が赤道板上でフラグモプラスト微小管の間に並び,これらが融合することにより形成されます。このように陸上植物の核分裂は
で特徴づけらます。
シャジクモ類(Chara, Nitella),コレオケーテ(Coleochaete)などMLS型鞭毛装置をもつ藻類のほか,アオミドロ(Spirogyra)やミカヅキモ(Closterium),ツヅミモ(Cosmarium)などの接合藻類も核分裂は開放型であり,フラグモプラスト微小管を使用して細胞板を形成します。接合藻類は生活環のなかで遊泳細胞を形成しないが,細胞分裂の性質に基づいてシャジク藻類と同じ仲間とされます。MLS型鞭毛装置をもつ糸状の緑色藻類のクレブソルミディウム(Klebsormidium)では,細胞板は形成されず,細胞膜の環状収縮によって細胞質分裂が行われるが,核分裂は開放型で紡錘体は終期まで残る点で,陸上植物型の分裂であるといえます。
これに対して,時計回り配列の交叉型鞭毛装置をもつクラミドモナス(Chlamydomonas)など淡水の緑藻類の多くでは,核分裂の時,核膜は消失しません。核膜を保ったまま(核膜残存),染色体は両極に分離し,娘核が形成されます。娘核が形成されるまでに中間紡錘体は崩壊して消失する(collapsing interzonal spindle)。紡錘体の消失に伴って2個の娘核は急速に接近して赤道面を挟んで向かい合うが,このとき,細胞の分裂軸に直角(つまり赤道面に水平)の方向に多数の微小管が出現します。この微小管をフラグモプラストに対してファイコプラスト(phycoplast)とよびます。ファイコプラストに沿って細胞膜の環状収縮や小胞の形成が起こって,やがて細胞の隔壁が形成されます。「閉鎖型の核分裂」と「ファイコプラストによる細胞質分裂」は時計回り配列の交叉型鞭毛装置をもつ緑色藻類に広く分布しています。淡水産緑藻の細胞分裂は
で特徴づけられます。
反時計回りの交叉型鞭毛装置をもつアオサなどの海産緑藻類も,淡水産の緑藻と同様に核分裂は閉鎖型です。核は核膜を保ったまま細長く伸びて,2個の娘核をつくります。中間紡錘体は分裂の終わり近くまで残り,その間に環状収縮が進行して細胞質が分裂します。細胞の隔壁の形成にフラグモプラストやファイコプラストは関与しません。海産緑藻類の細胞分裂は
で特徴づけられます。
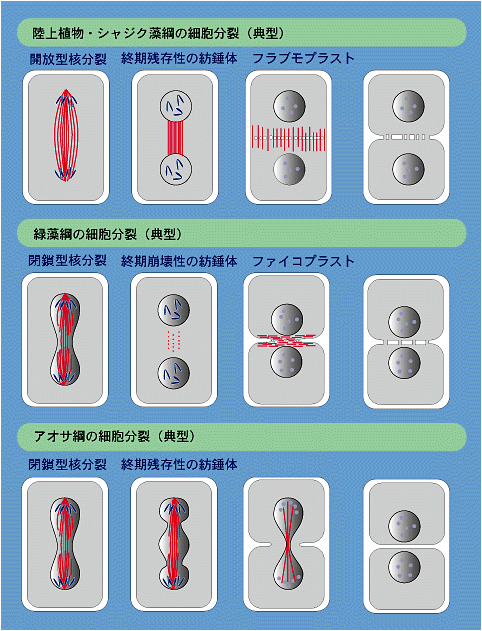
以上のように,緑色植物に系統の異なる3つの仲間が存在することは鞭毛装置と細胞分裂装置という細胞の活動に極めて重要な2つの構造によって支持されます。現在では,これらは綱の階級で分類されます。
シャジク藻綱は陸上植物に直接つながる系統群と考えられています。
細胞構造の比較から明らかになった分類群は生活環という側面でみても興味深いものです。緑藻綱のほとんどは淡水に生息し,栄養体が半数体であることが特徴です。これらは接合すると接合子が休眠し,発芽時に減数分裂を行って半数体に戻ります。したがって世代の交代はありません。シャジク藻綱の仲間も緑藻綱とほぼ同じ生活環を示します。これに対して,アオサ藻綱では栄養体が単相のもの,複相のもの,世代交代を行うものと行わないものなど,生活環は多様です。
このような性質は海という安定な環境とより大気の影響を受けやすい淡水の過酷な環境(乾燥や温度変化)に対応したもので,系統とのつながりを考えるのは必ずしも容易ではありません。
これら3つの大きなグループのほかに,プラシノ藻とよばれる緑色植物の一群があります。これは大部分が細胞壁をもたない単細胞遊泳性の種からなる小さなグループで,鞭毛と細胞の表面に鱗片をもち,鞭毛が細胞前端のくぼみから生じるなどの性質をもつことで,まとめられたグループです(鱗片は実際には上記の3つの系統のすべてに存在する。それぞれの綱のページを参照)。プラシノ藻のもつ多くの性質は緑色植物のなかで原始的と判断されることから,現在ではシャジク藻綱,緑藻綱,アオサ藻綱という緑色植物の3つの分類群の祖先群と考えられています。
{kind=link}
{kind=link}