![]()
![]()
![]()
![]()
![]()
ユーグレノゾア EUGLENOZOA
![]()
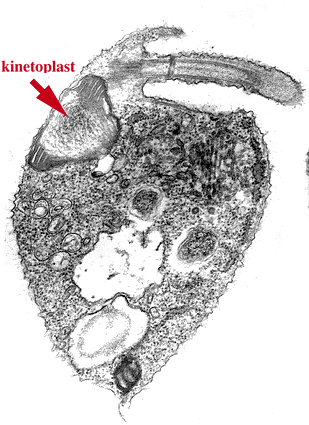
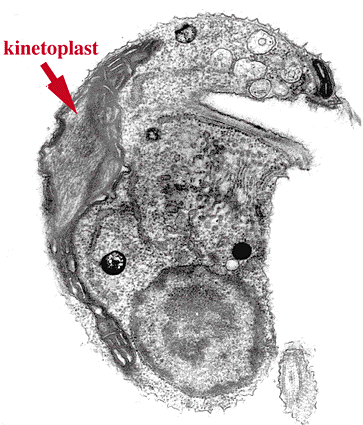
![]() キネトプラスト類は光学顕微鏡で識別できるミトコンドリアDNA 顆粒であるキネトプラスト(kinetoplast)をもつことでまとめられるなかまです。キネトプラストは最初に発見された核の外側にあるDNA(核外DNA)で,その発見は1924年にさかのぼります。葉緑体やミトコンドリアが独自のDNAをもつことが発見され,核外DNAの存在が一般に認められたのは1960年代のことです。キネトプラスト類のミトコンドリアDNAは多数のコピーをもち,凝集して存在するために古くから観察されていたのです。
キネトプラスト類は光学顕微鏡で識別できるミトコンドリアDNA 顆粒であるキネトプラスト(kinetoplast)をもつことでまとめられるなかまです。キネトプラストは最初に発見された核の外側にあるDNA(核外DNA)で,その発見は1924年にさかのぼります。葉緑体やミトコンドリアが独自のDNAをもつことが発見され,核外DNAの存在が一般に認められたのは1960年代のことです。キネトプラスト類のミトコンドリアDNAは多数のコピーをもち,凝集して存在するために古くから観察されていたのです。
トリパノソーマ類のすべてとボド類の一部は寄生性で,脊椎動物のすべての綱,甲殻類,昆虫,さらに植物にまで幅広い生物を宿主として寄生し,さまざまな病気を引き起こします。特にトリパノソーマ類のTrypanosomaによって引き起こされる眠り病 (sleeping sickness) は,およそ5000万人が感染しているといわれ,現在でも多くの人命を奪っている疾病です。LeishmaniaもTrypanosomaとともに多くの被害をもたらしています。
寄生性病原虫を中心とするこの鞭毛虫のなかまとユーグレナ植物の類似は古くから指摘されていたが,微細構造の比較研究および分子系統解析からその近縁性が明らかになってきました。
うちわ形のミトコンドリアクリステをもつ。 ミトコンドリアDNAは凝集して塊をつくっており,キネトプラストとよばれる。 | ||
  | ||
| 鞭毛軸糸に沿ってparaxial rodが存在する。 |
| |
| マスチゴネマをもつとき,ユーグレナと同様細い小毛である。 | ||
| 鞭毛装置 3個の鞭毛根をもつ。これらの起点はユーグレナ類のそれと一致しており,相同と考えられる。 |
| |
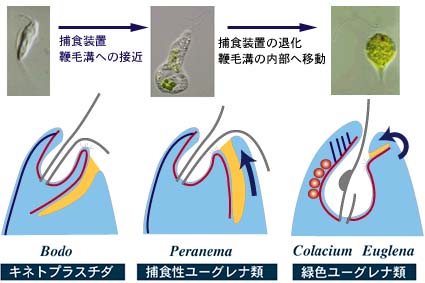
| キネトプラスト類のFd (dorsal fibrils)は細胞骨格を構成しており,一部のキネトプラスト類ではユーグレナと同様な複雑ならせん状の配列をもち,ユーグレナ類のペリクル微小管と相同と考えられる。mtr (reinforced microtubular root) は捕食装置(cytostome/cytopharynx) に伸長して付随することから,ユーグレナ類のverntral rootと相同と考えられる。また鞭毛根との位置関係から捕食装置もまた,互いに相同の構造と考えられる。 |
| |
| Entosiphonのような捕食生活をするユーグレナ類のほか,独立栄養で生活するEuglenaとColaciumでも退化した捕食装置がある。このことは,緑色のユーグレナ類が光合成能力を獲得する前は捕食によって生活する鞭毛虫であったことを示唆している。 | ||
| 核分裂は完全に閉鎖型であり,分裂期を通して核膜が残存する。したがって紡錘体は核内に形成される。中期赤道板は形成されない。 | ||
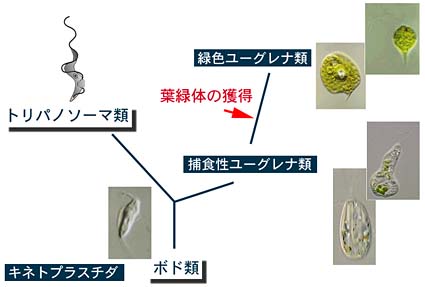
以上のような形態の類似から,現在ではユーグレナの祖先はボド類のような捕食性の鞭毛虫であり,そこから寄生性のトリパノソーマ類と自由生活で捕食性のユーグレナ類が分岐し,後者がやがて細胞共生によって葉緑体を獲得して光合成を行うユーグレナ類が出現したと考えられています。Astasiaのような吸収栄養で生活するユーグレナ類は二次的に葉緑体を失った仲間と考えられます。
分子情報もユーグレナ類とキネトプラスト類が単系統である(共通の祖先生物に由来する)ことを示しており,形態形質と分子形質が結論を相いに補強しています。共通の祖先に由来するユーグレナ類とキネトプラスト類はユーグレノゾア(Euglenozoa)の名前でひとつの生物群として認識されつつあります。
{kind=link}