|
研究内容
私の研究室では,神経細胞におけるシグナル伝達(シグナル・トランスダクション)
と細胞内情報処理の分子メカニズム解明することを目的に,主として脊椎動物の感覚細胞を用いて研究を行っている。研究方法として,パッチクランプ法や吸引電極法による電流記録などの電気生理学的方法を中心に,抗体を用いた免疫組織化学的方法,イオン画像解析法(イメージング)などを用いている。また必要に応じてコンピューターシミュレーションを取り入れたり,共同研究によってノックインマウスを用いた研究も開始した。
1. 視細胞における光シグナル伝達
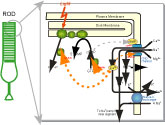 私たちは星明りのきわめて弱い光から太陽が降り注ぐ夏のビーチの光まで,広いダイナミックレンジの光を見ることができる。それは私たちの視覚系がさまざまなレベルで光の調節を行っているからである。視細胞中でもそのような調節のメカニズムが存在し,それは光シグナル伝達のプロセスの中で説明できる。 私たちは星明りのきわめて弱い光から太陽が降り注ぐ夏のビーチの光まで,広いダイナミックレンジの光を見ることができる。それは私たちの視覚系がさまざまなレベルで光の調節を行っているからである。視細胞中でもそのような調節のメカニズムが存在し,それは光シグナル伝達のプロセスの中で説明できる。
光が照射されると視細胞外節中ににある視物質(visual pigment)によってフォトン(光量子)が吸収され,視物質が活性化する。続いてトランスデューシン(Gタンパク質)が活性化し,さらにフォスフォジエステラーゼ(PDE)が活性化することによって,2次メッセンジャーであるcGMPが分解される。その結果,形質膜にあるCNGチャネルが閉じ,内向き電流が流れなくなることによって膜電位が減少する。これが視細胞における光応答である。CNGチャネルを通って流れ込むCa2+は,グアニル酸シクラーゼやロドプシンキナーゼを抑制し,cGMPのレベルを調節する。CNGチャネルから流れ込んだCa2+は,Na/Ca-K交換ポンプによって排出される。cGMPのレベルとCNGチャネルのオープニングの間にはCa2+を媒体とする負のフィードバックが存在し,これが視細胞における明順応の原因となっている (視細胞における光シグナル伝達のページ参照)。
私たちは現在,この光シグナル伝達の過程におけるCa2+やMg2+のはたらきについて,電気生理学的方法やイメージングのテクニックを使って研究を進めている。また,桿体(rod)と錐体(cone)では吸収波長が異なるだけでなく,光応答の感度やキネティクスが大きく違うが,その理由については不明である。それを明らかにする目的で,2004年の春から京都大学の七田研究室と共同で,遺伝子操作した視物質を持ったマウス(ノックインマウス)を作製し,アミノ酸配列の違いによる応答特性の変化を調べる実験を始める。
2. 嗅細胞におけるシグナル伝達
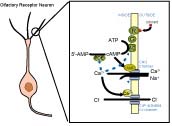 一説によると私たちが嗅ぎ分けられる匂いは10万種類と言われている。しかし現在知られているレセプター蛋白は約1000種類であり,1個の嗅細胞には1種類のレセプターが発現していると言われる。このように限られた数のレセプターによって多くの匂いを検出するには,嗅細胞の内部や嗅覚系においてさまざまな情報処理が行われると考えられる。 一説によると私たちが嗅ぎ分けられる匂いは10万種類と言われている。しかし現在知られているレセプター蛋白は約1000種類であり,1個の嗅細胞には1種類のレセプターが発現していると言われる。このように限られた数のレセプターによって多くの匂いを検出するには,嗅細胞の内部や嗅覚系においてさまざまな情報処理が行われると考えられる。
嗅細胞におけるシグナル伝達については次のことが明らかになっている。匂い分子(odorannt)がレセプター蛋白によって受容されると,Gタンパク質を介してアデニル酸シクラーゼが活性化され,2次メッセンジャーであるcAMPが合成される。その結果,形質膜にあるCNGチャネルが開き,内向き電流が生じることによって膜電位が上昇する。CNGチャネルを通って流れ込んだCa2+は,形質膜にあるCa2+依存性Cl-チャネルを活性化し,Cl-が外向きに流れることによって内向き電流が生じ,膜電位がさらに上昇する。このようにして匂い応答が生じる (嗅細胞におけるシグナル伝達のページ参照)。
最近の研究によれば,ある種の匂い物質が電気 的信号を抑制することが報告されれいおり,我々もその現象を嗅細胞レベルで確認した。現在は,この"抑制性匂い応答"のメカニズムを解明する研究を進めるとともに,嗅細胞レベルで,異なった種類の匂い同士の相互作用とその分子メカニズムを調べる研究,嗅細胞におけるアミノ酸応答の研究などを行っている。
私の研究室では,主として一般生理学的な観点から研究を進めているが,最近比較生理学的な観点から,板鰓類(サメ)の嗅覚系についての生理学的研究も行い,感覚メカニズムの多様性についても調べている。
3. 味細胞におけるシグナル伝達
味は甘味,塩味,酸味,苦味,旨味の5種類に大別される。味細胞におけるシグナル伝達のメカニズムは,味の種類によって大きく異なることが知られており,その詳細は明らかになっていない。私たちはカエル味細胞を用いて,シグナル伝達の分子メカニズムの研究を行っている。
|