![]()
高等植物における形態形成現象のうち最も劇的な変化の一つである胚発生現象に着目し、生化学的・分子遺伝学的視点から研究を行なっている。
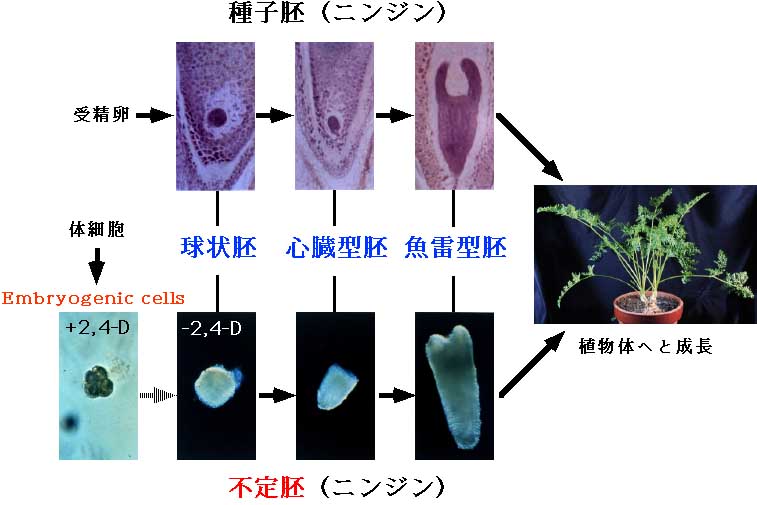
種子胚発生は胚珠内で進行するため、その解析が困難である。一方、不定胚形成系においては、球状胚、心臓型胚、魚雷型胚というように種子胚と同様の形態変化を経て幼植物体が形成される。特に、ニンジンの不定胚形成系は、培養により均一な胚を大量に調整することが可能であるため、種子胚発生のモデル系として広く利用されている。本研究室では、ニンジンの不定胚形成系を用いて高等植物の胚発生に関わる様々な生理現象の解明を目指している(不定胚形成のモデル)。
![]() 胚発生
胚発生
高等植物は一度決定した組織が脱分化を起こし再び完全な植物体に再生する能力、分化全能性を有する。なかでも、体細胞から生じる不定胚形成現象はこの分化全能性を実証する最も重要な現象である。本研究室では不定胚形成について、体細胞が胚発生を開始する機構を解析している。
不定胚の誘導は通常植物ホルモンの一種であるオーキシン(2,4-D)を用いて行なわれ、その誘導は2,4-Dの添加・除去によって行なうことができる。そのためこれまでの定説では、オーキシンが不定胚形成を誘導するとされてきた。オーキシンを用いない不定胚誘導系(ストレス不定胚誘導系)を用いた研究から、体細胞が不定胚を形成する現象はオーキシンの作用に加え、体細胞にストレスが与えられることによる生体内アブシジン酸(ABA)含量の増加が、胚発生誘起に関係している可能性が示唆された。
また、これらの不定胚誘導系において、胚発生能力獲得時に共通して発現するタンパク質(ECP)遺伝子群の同定・単離にも成功した。これらのECP遺伝子のプロモーター解析による発現制御機構の解析を行っている。一方、これらのECP遺伝子の発現を制御する転写制御因子として、C-ABI3遺伝子の単離に成功している。C-ABI3の発現制御機構の解析により、胚発生の初期を制御する胚決定因子を解明が期待される。また、C-ABI3と同様に、胚発生に関わる転写因子として、C-LEC1, C-FUSの解析も進行している(胚発生機構のモデル)。
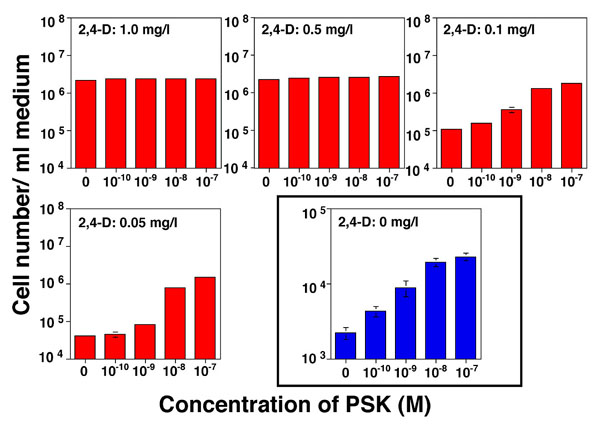
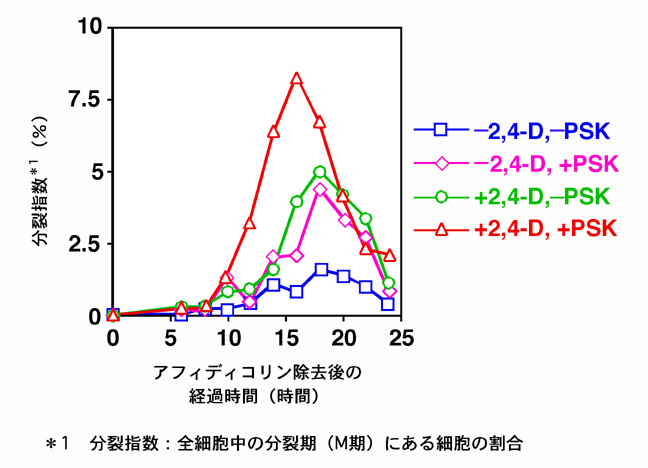
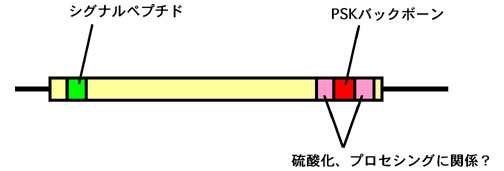
動物においては、ペプチド性の増殖因子は非常に一般的であるが、植物においては、ほとんど知られていない。1996年にアスパラガス葉肉細胞培養液より単離されたファイトスルフォカイン(PSK)は、世界で初めて植物より単離されたペプチド性の細胞増殖因子である。PSKは、低細胞密度における細胞分裂促進作用を持つことが示されてきているが、その細胞分裂活性化のメカニズムは明らかにされていない。そこで、PSKが、植物個体内でどのようなメカニズムにより働いているのかについて、細胞レベルないし個体レベルで明らかにすることを目的として研究を進めている。細胞レベルの研究については、ニンジンのnon-embryogeniccell(NC)と呼ばれる、細胞増殖のみを行う細胞培養系を用いて、これまでに、低濃度の2,4-D存在下でのみPSKが増殖に影響を与えることを明らかにしている。また、同じNCを用いた同調培養系において、PSkの作用には細胞分裂活性と細胞周期進行促進の2つの効果があることも示された。一方、個体レベルの研究では、個体中でのPSKの存在部位について、PSKそのもの、またはその前駆体の発現部位を調べていくとともに、すでにイネにおいて単離されているPSK前駆体遺伝子を用いて過剰発現ないし発現抑制した場合の形態などの変化を解析している。またイネなどのタギングラインを用いて、遺伝子破壊株における表現型の解析も行っている。
![]() 胚発生阻害物質
胚発生阻害物質
ニンジンの不定胚誘導系では、細胞を高密度下で培養した場合、不定胚への分化・発達において阻害効果が観察される。当研究室では、この原因物質のひとつが既に単離され、p-hydroxybenzylalcohol(pHBA)であることが確認されている。このpHBAは不定胚だけではなく、ニンジンの未熟種子にも存在し、胚発生の際に特異的に合成され、pHBAは胚発生の特に初期の細胞分裂を阻害することが明らかとなった。現在、種々の植物におけるpHBAの効果の普遍性、生合成系、生理的な存在意義などの解明を進めており、pHBAによる阻害メカニズムの解明により、胚発生初期の特殊な細胞分裂の制御機構が明らかとなると予想される。
![]() 乾燥耐性
乾燥耐性
不定胚は種子胚と同じ様な形態変化を示すが、受精胚は成熟後期に乾燥して休眠するのに対して、不定胚は乾燥させると死滅し、乾燥耐性がみられない。種子では成熟後期には植物ホルモン、アブシジン酸(ABA)が増加するのに対して、不定胚ではほとんどABAは存在しない。このABAが存在しないことが、不定胚が乾燥耐性を持たない理由と考えられた。実際に、不定胚にABA処理を施すことにより、極めて強い乾燥耐性を不定胚へ付与できることが明らかとなった(ABA処理による乾燥耐性獲得)。
現在、乾燥耐性誘導時に転写・翻訳される各種遺伝子の転写制御におけるこれら遺伝子の機能解明を進めるとともに、自身の転写制御因子について同定・単離を進めている。
![]() シロイヌナズナを用いた研究
シロイヌナズナを用いた研究
ニンジンは二年生植物であること・ゲノムが大きいことから、突然変異体の解析や遺伝学的解析には適していない。一方、シロイヌナズナはこのような解析に適しており、高等植物のモデル植物として広く利用されている。そこで、シロイヌナズナを用いて、胚形成関連遺伝子(ECPs,
ABI3, LEC, FUS)の突然変異体の解析や遺伝学的解析を行っている。また、当研究室において、シロイヌナズナの不定胚誘導系を開発しており、この不定胚誘導系を用いた胚発生機構の解析が期待される。
以上のような研究を通じ、受精卵や胚を構成する細胞が胚であることを規定する分子機構を解明、胚発生を通じて高等植物の形態形成現象を究明したいと考えている。
![]()
![]()
{kind=link}
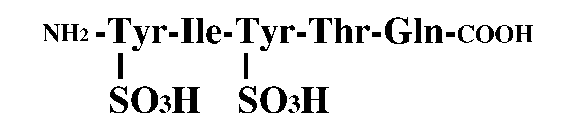{kind=link}
{kind=link}
{kind=link}
{kind=link}