
図1. 左から: ツリガネニンジン (キキョウ科) 、セイヨウアブラナ (アブラナ科) 、オオアワガエリ (イネ科) 、シキミ (シキミ科) 、ファレノプシス (ラン科).
|
|
花 (flower) は、生殖器をつけた葉とその周囲の葉が変化してできた被子植物に特有の器官である。花は雄しべのつくった花粉と雌しべの中の胚嚢が合体して次世代の植物体である種子をつくるという直接的な機能のほかに、それを効率的に行いかつ保護するという機能を果たすため、萼片や花弁などの花被片をもっている。このような生物学的な機能に適応し、そして植物がたどってきた進化の道筋の跡を残しているために、花は基本プランを共有するとともに非常な形態的多様性を示す (図1)。
広い意味で言えば、裸子植物やシダ植物も"花" (生殖器をつけた葉の変形物) をもっているということができる (図2)。しかし、これらの"花"は被子植物の花とは構造的に大きく異なり、直接的に比較するのは困難である。一般的には花とは被子植物の器官に限られる。
被子植物の花は基本的に外側から萼片、花弁、雄しべ、雌しべが輪状に集まってできている (図3)。これらの花要素はすべて葉が変化してできたものであり、花葉 (floral leaf) とよばれる。これらの要素が葉から変化したものであることは、近年の分子生物学的な研究に基づく実験からも確かめられている (ABCモデル) 。
|

図2. 左から: アカマツ (マツ科) の雄性球花、ソテツ (ソテツ科) の雌性胞子葉群、マンネンスギ (ヒカゲノカズラ科) の胞子嚢穂.
|
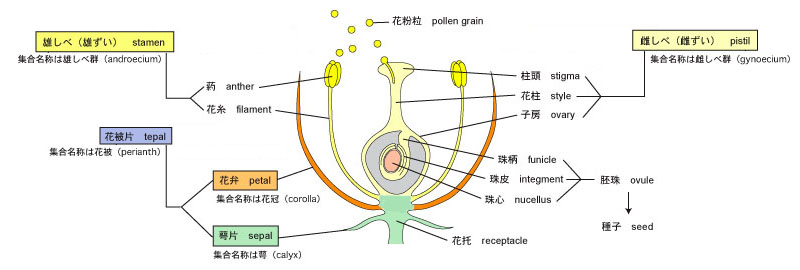
図3. 被子植物の花の模式図
|
 萼片と萼 萼片と萼
花の最外輪にある花葉で、その内側の要素 (花弁) とは質が異なる場合、その個々の要素を萼片 (sepal) 、要素をまとめて萼 (calyx) とよぶ (図4)。ふつう萼は花の中で葉の特徴を最もよく残しており、他の要素を保護する役割を果たしているが、トリカブト属 (キンポウゲ科) のように花弁の代わりに目立つ色・形をしていることもある。萼片は互いに離生しているものもあるし、互いに合着して萼筒 (calyx tube) を形成していることもある (図4左)。
 萼片と萼について 萼片と萼について
|

図4. 左:ハナトラノオ (シソ科) では5個の萼片が合着して萼筒を形成している. 右:ヘビイチゴ (バラ科) には大きな萼と副萼がある.
|
花弁と花冠
萼の内側、雄しべの外側にある花葉で、萼とは質が異なる場合、個々の要素を花弁 (petal) 、要素をまとめて花冠 (corolla) とよぶ (図5)。花弁とは多くの場合、いわゆる"花びら"のことである。花冠は内側の雄しべや雌しべを保護すると同時に、昆虫の誘因など効率的な花粉媒介を助ける働きを果たしている。花弁が離生している花を離弁花 (choripetalous flower) (図5左)、互いに合着している花を合弁花 (sympetalous flower) (図5右) という。
花弁と花冠について
|

図5. 左:アケボノスミレ (スミレ科) の花は5個の花弁が左右相称にならぶ離弁花である. 右:ヤマホタルブクロ (キキョウ科) では5個の花弁が合着して鐘状の花冠を形成する.
|
花被片と花披
萼片と花弁をあわせて花被片 (tepal) とよび、その全体名称は花披 (perianth) という。特にユリ (ユリ科) やモクレン (モクレン科) など萼片と花弁の分化が不完全な花に用いられる (図6)。花被が内外2輪になっている場合、外側を外花被 (outer perianth) 、内側を内花被 (inner perianth) という。進化的には内外の花被片が分化していない状態が原始的だと考えられる。
|

図6. 左:ハクモクレン (モクレン科) では花被片が3個ずつ3輪に配置している. 右:スカシユリ (ユリ科) では外花被片3個、内花被片3個がある.
|
花托と花軸
花葉をつけている茎の先端が変化した部分を花托 (tarus) とよぶ。子房周囲や子房下位の花では、ふつう花托が筒状になって子房を包んでおり、この筒を花托筒 (hypanthium) という。また子房基部 (または花柱基部) の花托が肥大したものは花盤 (disc) とよばれる。ハス (ハス科) ではさらに花托が発達して雌しべ全体を包み込んでしまっている (図9左)。またタイサンボク (モクレン科) のように花托が上下に長い場合は、花軸 (floral axis, rachis) とよぶ (図9右)。フウロソウ属 (フウロソウ科) では花後に雌しべ内 (心皮の間) を花軸が伸びるが、これを心皮間柱 (carpophore) という。またナデシコ属 (ナデシコ科) では萼と花冠の間の花軸が伸び、花被間柱 (子房柄 gynophore) とよばれる。
|

図9. 左:ハス (ハス科) の花托は大きく発達して複数の雌しべを包んでいる. 右:タイサンボク (モクレン科) の花托は前後に長くなり花軸となる.
|
花柄と苞
1つの花を支える柄のことを花柄 (pedicel) とよび (図10左)、花の基部にあって他の葉とは質が異なる葉のことを小苞葉または苞葉 (bracteole) とよぶ (図10右)。特に小さな花が集合している場合、個々の花を小花 (floret) 、柄を小花柄 (pedicelet) ということがある。
|

図10. 左:ツリガネニンジン (キキョウ科) の花は長い花柄の先についている. 右:ミズギボウシ (ユリ科) の花の基部には小さな苞がある.
|
花床と花梗、総苞
キク科のように多数の花が集合して1つのまとまりをつくる場合、花がついている平面的に広がった部分を花床 (receptacle) とよぶ (図11左)。また複数の花をつける共通の枝を花梗 (peduncle) という (図11右)。複数の花 (花序) の根元にあって他とは異なる葉のことを総苞片 (involucral segment) とよび、集合名称として総苞 (involucre) という。タンポポなどキク科の花で萼のように見えるものは総苞である (図11左)。
|

図11. 左:セイヨウタンポポ (キク科) は多数の舌状花が花床についており、その根元には多数の総苞片がある. 右:アセビ (ツツジ科) は花梗に多数の花がついている.
|




