|
植物 (plants) とは何だろうか? ふつう植物とは、水と二酸化炭素、光エネルギーから酸素と糖を合成する光合成、つまり酸素発生型光合成 (oxygenic photosynthesis) を行う生物と定義される (図1)。私たちに身近なタンポポやイネ、マツ、シダ、コケなどと共に、海にすむアオノリやテングサ、コンブのような海藻や、クロレラ (Chlorella) 、クラミドモナス (Chlamydomonas) 、ミドリムシ (Euglena) 、珪藻のような単細胞生物までもが植物であるといえる (図2)。植物は水圏から陸上まで、生態系の生産者 (producer) として欠くことのできない役割を担っている。ただし植物の中には、二次的に光合成能を失った生物 (寄生植物、腐生植物など) も含まれる (図5)。
|
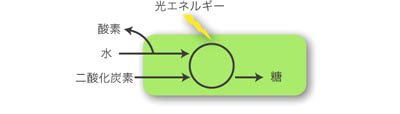
図1. 酸素発生型光合成の模式図
|
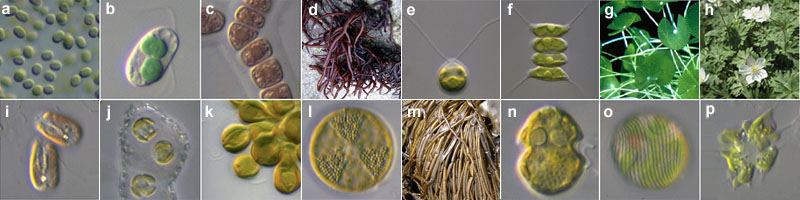
図2. さまざまな植物 a. Aphanocapsa (シアノバクテリア). b. Cyanophora (灰色植物). c. Asterocytis (紅色植物). d. フクロフノリ (紅色植物). e. Chlamydomonas (緑色植物). f. Desmodesmus (緑色植物). g. カサノリ (緑色植物). h. ニリンソウ (緑色植物). i. Rhodomonas (クリプト植物). j. Umbiricosphaera (ハプト植物). k. Synura (不等毛植物). l. Actynoptycus (不等毛植物). m. カヤモノリ (不等毛植物). n. Heterocapsa (渦鞭毛植物). o. Lepocinclis (ユーグレナ植物). p. Lotharella? (クロララクニオン植物).
|
|
では、この意味での植物とはひとまとまりの、つまり単系統 (monophyly) の生物群なのだろうか? 答えは否である。酸素発生型光合成は最初、原核生物において誕生した。この子孫がアオコなどの藍色植物 (cyanophytes) 、つまりシアノバクテリア (藍色細菌 cyanobacteria) である (図2a)。この藍色植物 (藍藻) は、ある真核生物に取り込まれ、共生関係 (一次共生 primry endosymbiosis) を築き上げ、葉緑体 (chloroplast) へと変化した。この一次共生という現象によって、酸素発生型光合成という機能が真核生物に伝えられたのだ。その子孫が灰色植物 (図2b)・紅色植物 (テングサやオゴノリなど) (図2c-d)・緑色植物 (イネやクロレラなど) (図2e-h) である。この3植物群は一次植物 (primary plasts) ともよばれ、また系統分類学的にはこの3植物群のみを狭義の植物 (kingdom Plantae) とする意見もある。さらにこの狭義の植物が二次共生 (secondary endosymbiosis) という現象によってさまざまな真核生物に取り込まれ、葉緑体となることで新たな植物が生じた。その結果生まれたのがミドリムシや珪藻、ワカメなどの植物 (二次植物 secondary plants) である (図2i-p)。つまり酸素発生型光合成は、共生という現象を通じてさまざまな生物が獲得した機能であり、それによって定義される植物というまとまりは多系統 (polyphyly) である (図3) 。
しかし私たちがふつう植物として認識しているのは、陸上にすむコケやシダ、裸子植物 (イチョウ、ソテツ、マツなど) 、被子植物 (スイレン、イネ、ダイコン、キクなど) である。系統分類学的にも、植物という用語をこれらの生物群に限る場合がある。これらの植物は陸上植物 (land plants) とよばれ、共通の祖先をもつ単系統群である。これに対して、陸上植物以外の植物は藻類 (algae) と総称される。陸上植物は陸上生態系のほとんどの生産をまかなっていると同時に、食用 (コショウ、トウモロコシ、コムギ、イネ、リンゴ、ジャガイモ、レタスなど) 、材料用 (ヒノキ、ワタ、ミツマタなど) 、観賞用 (ユリ、ラン、ナデシコ、サクラ、バラ、キクなど) として人間生活に密接に関わっている。以下このリソースで植物として扱うのは基本的に陸上植物、特に維管束植物のことである。
|

図3. 植物 (酸素発生型光合成生物) を中心とした生物の系統。色の付いた系統は植物を、矢印は共生を示す。共生の回数はもっと少なかったかもしれない。
|
 植物と動物 植物と動物
古典的には、植物は動物とともに生物界を大きく2分する単位であった (2界説)。しかし系統的に考えると、先に述べたように植物自体が多系統群であり、動物 (捕食栄養性生物) もまた多系統群である。したがって植物と動物をはっきりと区分する形質は存在しない。
動物は動き、植物は動かないという分け方は直感的であるが、動物でもカイメン (海綿動物) やサンゴ (刺胞動物) は一度固着すると移動しない (図4) 。また植物でも、藻類のなかには鞭毛運動によって動くものが少なくない (クラミドモナスやミドリムシなど) (図2)。陸上植物は、個体が自力で移動することはないが、多くの花や葉が夜には閉じるなどゆっくりとではあるが運動をすることがよく知られている。またオジギソウ (マメ科) の葉が接触刺激によって閉じることや、ハエトリソウ (モウセンゴケ科) の葉が昆虫を捕らえるときに閉じる反応は素早いものであり、動物の運動速度に劣らない。
植物は光合成を行う生産者(光独立栄養性)であり、動物は補食する消費者(従属栄養性)であるという生態学的観点からの区分はわかりやすいが、これにも例外が多い。例えば寄生植物のラフレシア (ラフレシア科) やヤセウツボ (ハマウツボ科) 、腐生植物のギンリョウソウ (ツツジ科) やオニノヤガラ (ラン科) は光合成能を欠く (図5)。しかし寄生性や腐生性の植物でも,細胞小器官としての葉緑体 (色素体) は残しており,このような光合成能を失った色素体を白色体 (leucoplast) という。また動物の中には藻類を共生させて生産者として生きるものが少なくない (図4)。特に熱帯浅海域におけるサンゴは渦鞭毛植物を共生させており、見かけ上は生産者として重要な地位を占めている。
しかし植物と動物をそれぞれ我々に身近な陸上植物と多細胞動物 (後生動物) に限ると,さまざまな相違点がある。特に細胞レベルではその違いは明瞭で,例外なく分けることができる (表1)。
| |
陸上植物 |
後生動物 |
| 生活様式 |
生産者* |
消費者** |
| 運動 |
ゆっくり* |
すばやい* |
| 色素体 |
あり |
なし |
| 中心体 |
なし |
あり |
| 細胞壁 |
あり |
なし |
| 核分裂 |
開放型 |
閉鎖型 |
| 細胞質分裂 |
遠心的(フラグモプラスト) |
求心的(収縮環) |
| 細胞間の連絡 |
原形質連絡 |
細胞膜が直接接する |
表1. 陸上植物と後生動物の比較 (*例外あり、**藻類や化学合成細菌を共生させて生産者となるものもある)
|

図4. "植物的"な動物. 左:ミズタマサンゴ (刺胞動物) は固着すると動かない。右:オヨギイソギンチャク (刺胞動物) は渦鞭毛藻 (Symbiodinium) を共生させて光合成を行う。

図5. 光合成能を失った植物. 左:ヤセウツボ (ハマウツボ科) は寄生植物であり、ムラサキツメクサ (マメ科) などから栄養を吸収している。右:ギンリョウソウ (ツツジ科) は腐生植物であり、根についた菌類から栄養を吸収している。
|
陸上植物
陸上植物はコケ植物と維管束植物からなる。維管束植物にはシダ植物と種子植物が含まれ、種子植物は裸子植物と被子植物からなる。陸上植物にのなかには、アカウキクサ (ウラボシ綱) やマツモ (マツモ科) 、アマモ (アマモ科) など水生のものも知られるが、これらは陸上から二次的に水に戻ったものであり、陸上植物に含められる (図6)。陸上植物とはその共通祖先に由来する生物群の名称であり、陸棲であっても藻類は陸上植物には含めない。
陸上植物に共通する最も重要な特徴は、多細胞性の (雌性) 生殖器の中に幼い胞子体、つまり胚 (embryo) を形成することである。そのため陸上植物は有胚植物 (embryophytes) とよばれることがある。またほとんどの藻類とは異なり、陸上植物は多細胞性の造卵器 (archegonium) をもつので、造卵器植物 (Archegoniatae) ともいう。ただし種子植物の造卵器は極めて退化的であるため、狭義の造卵器植物からは除かれることもある。
|

図6. 水生の陸上植物. 左: ウミショウブ (トチカガミ科). 右: ウキクサ (サトイモ科).
|
コケ植物
コケ植物 (bryophytes) には苔類、蘚類、ツノゴケ類の3グループがある (図7)。コケ植物は本体が配偶体 (単相) であり、配偶子 (卵と精子) を形成する。有性生殖は卵と精子 (2本鞭毛性) の受精によってなされ、その結果生じた胞子体 (複相) は配偶体に寄生したまま成長し、減数分裂によって胞子を形成する (図8)。植物体のつくりは単純で、組織の分化は不完全であり、維管束を欠く。これらの特徴は陸上植物における原始的な状態を示すものである (共有原始形質) 。コケ植物は原始的な陸上植物の寄せ集めであって単系統群ではない。近年の分子系統学的研究によって、陸上植物の中で苔類がもっとも初期に分岐し、ツノゴケ類が維管束植物の姉妹群であることが示唆されている。
 コケ植物について コケ植物について
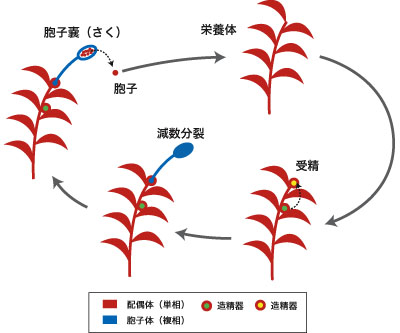
図8. コケ植物の生活環. コケ植物の栄養体は配偶体(単相)であり、そこに造卵器・造精器が形成、受精卵は配偶体についたまま胞子体へと成長する。胞子体の胞子嚢(さく)では減数分裂によって胞子が形成され、これが新しい配偶体になる。
|

図7. さまざまなコケ植物. 上左:ゼニゴケ (苔植物門). 中左:オオミズゴケ (蘚植物門). 中右:? (蘚植物門). 下:ナガサキツノゴケ? (ツノゴケ植物門).
|
維管束植物
維管束植物 (vascular plants, tracheophytes) にはシダ植物と種子植物が含まれる。維管束植物の本体は胞子体 (複相) であり、配偶体 (単相) は小形でときに胞子体に寄生した状態にある。
コケ植物にくらべて体のつくりは複雑で、ふつう根・茎・葉からなる。また組織の分化が見られ、水や無機栄養、同化産物の通道組織として維管束が分化している。維管束には植物体の機械的支持の役割もあり、ときに100 m以上にもなる大きな体を支えている。維管束植物は共通祖先に由来する単系統群であり、おそらく古生代シルル紀に出現した。
|
|
シダ植物
シダ植物 (pteridophytes) にはヒカゲノカズラ綱、トクサ綱、マツバラン綱、リュウビンタイ綱、ウラボシ綱の5グループがある (図10)。シダ植物は本体が胞子体 (複相) であり、減数分裂によって胞子を形成する。胞子は発芽して前葉体とよばれる配偶体 (単相) になり、ふつう胞子体とは独立して生活する。前葉体上に卵と精子 (2本または多鞭毛性) がつくられ、その受精によって新たな胞子体が形成される (図11)。植物体は基本的に根・茎・葉からなるが、その葉はグループによって起源が異なると考えられている。シダ植物の特徴は維管束植物における共有原始形質であり、シダ植物は単系統群ではない。シダ植物のなかではヒカゲノカズラ綱が最も初期に分岐し、他の4グループは単系統群をなして種子植物の姉妹群であると考えられている。
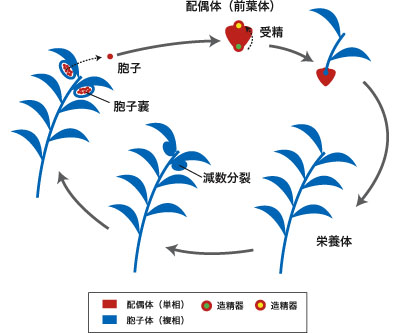
図11. シダ植物の生活環. シダ植物の栄養体は胞子体(複相)であり、減数分裂によって胞子が形成される。胞子は発芽して配偶体(前葉体)となり、そこに造卵器・造精器が形成される。受精卵は配偶体についたまま胞子体へと成長するが、栄養的には配偶体とは独立している。
|

図10. シダ植物のいろいろ. 上左:コンテリクラマゴケ (ヒカゲノカズラ綱). 上右:ミズニラ (ヒカゲノカズラ綱). 中上左:マツバラン (マツバラン綱). 中上右:ナツノハナワラビ (マツバラン綱). 中下左:Equisetum myriochaetum (トクサ綱). 中下右:リュウビンタイ (リュウビンタイ綱). 下左:Blechnum gibbum (ウラボシ綱). 下右:デンジソウ (ウラボシ綱).
|
種子植物
種子植物 (seed plants, spermatophytes) には裸子植物と被子植物がある。種子植物の本体は胞子体 (複相) であり、減数分裂によって大胞子と小胞子を形成する。大胞子は胚珠という構造の中に形成され、そこで発芽して雌性配偶体 (胚嚢) になる。小胞子は花粉粒となり、風や昆虫によって胚珠へと運ばれる(花粉媒介について)。花粉は発芽して雄性配偶子である精細胞または精子を送り込み、胚珠の中にできた雌性配偶体の卵細胞と受精する。胚珠は受精卵から発生した胚を包んだまま成熟して種子となり、散布されて生育地を広げる。種子植物の体も根・茎・葉からなり、大きさ数 mm のものから100 m 以上に達するものまである。種子植物は共通祖先をもつ単系統群であり、古生代デボン紀後期に出現したと思われる。
|
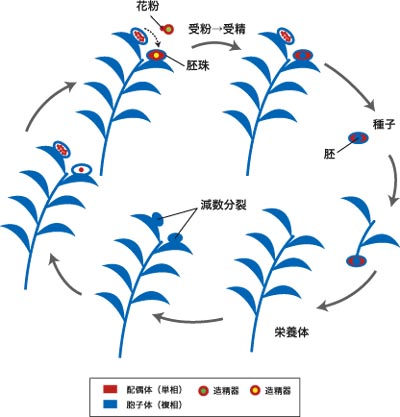
図12. 種子植物の生活環. 種子植物の栄養体は胞子体(複相)であり、減数分裂によって大胞子・小胞子が形成される。大胞子は胚珠内で形成され、そのまま雌性配偶体へと成長する。小胞子は花粉粒となり、雄性配偶体として精子または精細胞を形成する。
|
裸子植物
裸子植物 (gymnosperms) は古生代から中生代にかけて繁栄したが、現生の裸子植物としてはイチョウ、ソテツ類、球果類 (マツ、スギ、ヒノキなど) 、グネツム類の4グループのみが知られている (このうちグネツム類は特殊化した球果類かもしれない) (図11)。イチョウやソテツ類では、花粉粒が発芽すると多鞭毛性の精子がつくられるが、他では鞭毛を欠く精細胞がつくられる。胚珠内の雌性配偶体 (胚嚢) は多数の細胞からなり、数個の卵細胞が形成される。精細胞または精子は卵細胞と受精して胚になり、胚珠は種子になる。胚珠は裸出しているため (裸子の名の由来) 、種子はむき出しになる。これらの特徴は種子植物における原始形質であり、化石種を含めれば裸子植物は非単系統群であると思われるが、現生の裸子植物は単系統群であるらしい。
|

図11. 裸子植物の4つのグループ 左上:ソテツ (ソテツ目) 右上:イチョウ (イチョウ目) 左下:アカマツ (球果目) 右下:ウェルウィッチア (奇想天外) (グネツム目)
|
被子植物
スイレン、モクレン、コショウ、ミズバショウ、ヤシ、イネ、ケシ、クリ、カーネーション、バラ、タンポポなど我々に身近な植物のほとんどは被子植物 (angiosperms) である。生殖器官として花被片 (萼片と花弁) ・雄しべ・雌しべからなる花をつくる (図12) 。雄しべで花粉粒が、雌しべの中で胚珠がつくられる。雌性配偶体である胚嚢は胚珠の中につくられるが、裸子植物の雌性配偶体にくらべて極度に単純化しており、ふつう8核7細胞しかない。花粉は雌しべの先端 (柱頭) について発芽し、2個の精細胞が胚嚢へ送り込まれる。2個の精細胞のうち、1つは卵細胞と、もう1つは2個の極核をもつ中央細胞と受精する。このように2つの受精が起こることを重複受精といい、被子植物の重要な特徴である。胚珠は次世代の個体〔胚〕を含んだまま成熟して種子になるが、胚珠は雌しべ (の子房) に包まれており (被子の名の由来) 、包んでいる子房は成熟して果実になる。被子植物は共通祖先をもつ単系統群であり、おそらく中生代ジュラ紀ごろに出現した。
|

図12. いろいろな被子植物の花 左上:スイレンの園芸品種 'Star of Siam' (スイレン科) 右上:シダレザクラ (バラ科) 左下:イブキジャコウソウ (シソ科) 右下:サギソウ (ラン科)
|


